1998 में, आग और मेलो की रिपोर्ट है कि डबल असहाय शाही सेना (dsRNA) Caenorhabditis एलिगेंस 1 में जीन समारोह के निषेध के लिए प्रेरित कर सकते हैं। DsRNA से शुरू हो रहा यह प्रतिक्रिया शाही सेना हस्तक्षेप (आरएनएआई) नामित किया गया था, और जैसे आरएनएआई की मध्यस्थता जीन मुंह बंद जानवरों, पौधों, और कवक 2-7 में संरक्षित करने की सूचना मिली थी। पौधों और कुछ पशुओं में, आरएनएआई कार्यों प्रणालीबद्ध, जिसका अर्थ है प्रभाव अन्य कोशिकाओं / ऊतकों जहां dsRNA सीधे नहीं शुरू की है (8-10 में समीक्षा) में फैल सकता है। वैज्ञानिकों ने ब्याज की जीन को लक्षित करने के dsRNAs डिजाइन द्वारा इस अंतर्जात सेलुलर आरएनएआई प्रतिक्रिया का इस्तेमाल किया है, जिससे सीधे जीनोम (11-14 में समीक्षा) से छेड़छाड़ के बिना जीन समारोह नीचे दस्तक।
आरएनएआई निम्नलिखित लाभ के कारण कार्यात्मक अध्ययन के लिए एक शक्तिशाली उपकरण है। पहला, यहां तक कि कम से कम जीन अनुक्रम जानकारी के साथ एक जीन आरएनएआई का उपयोग कर निशाना बनाया जा सकता है। यह अनुसूचित जनजाति के लिए विशेष रूप से महत्वपूर्ण हैगैर-मॉडल जीवों जीनोमिक या transcriptomic डेटा की कमी के udies। दूसरा, जीवों जहां आरएनएआई प्रतिक्रिया मजबूती के साथ प्रणालीगत है, आरएनएआई की मध्यस्थता जीन पछाड़ना लगभग किसी भी विकास के चरण में किया जा सकता है। यह सुविधा pleiotropic जीन के समारोह के अध्ययन के लिए बहुत उपयोगी है। तीसरा, कुछ मामलों में, आरएनएआई प्रभाव gonads और संतान, ऐसा है कि phenotypes संतानों 15,16 में मनाया जाता है में फैल गया। यह घटना, माता पिता का आरएनएआई (pRNAi) के रूप में जाना जाता है, के रूप में एक ही इंजेक्शन माता-पिता द्वारा उत्पादित कई संतानों अंडे के प्रत्यक्ष हेरफेर के बिना जांच की जा सकती भ्रूण के विकास को प्रभावित जीनों के लिए विशेष रूप से फायदेमंद है। इन कारणों के लिए, pRNAi पसंद की विधि है। हालांकि, अगर pRNAi अप्रभावी है, oogenesis के लिए आवश्यक जीन के लिए उदाहरण के लिए, तो भ्रूण आरएनएआई (eRNAi) का इस्तेमाल किया जाना चाहिए। चौथा, आरएनएआई मजबूत दोषों को कमजोर उत्पादन करने के लिए है कि एक allelic श्रृंखला dsRNA की राशि वितरित एक सीमा से अधिक अलग किया जा सकता है के बराबर उत्पन्न करने के लिए इस्तेमाल किया जा सकता। phenotypes के इस तरह के एक उन्नयन जीन समारोह को समझने जब जीन एक जटिल प्रक्रिया है और / या समारोह का पूरा नुकसान में शामिल है घातक है के लिए सहायक हो सकता है। पांचवां, dsRNA के वितरण के लिए विशेष रूप से मजबूत प्रणालीगत आरएनएआई की प्रतिक्रियाएं दिखा पशुओं में, आम तौर पर आसान है और संभव है। dsRNA microinjection 1,5, खिला / घूस 17,18, भिगोने, 19,20 और वायरस / बैक्टीरिया की मध्यस्थता वितरण 21,22 द्वारा शुरू की जा सकती है। छठी, कुछ जीन लक्ष्यीकरण / संपादन विधियों के विपरीत, वहाँ उत्परिवर्तन ले जीवों के लिए स्क्रीन करने के लिए या आनुवंशिक पार जब आरएनएआई का उपयोग कर समयुग्मज उत्पन्न करने के लिए बाहर ले जाने की कोई जरूरत नहीं है। इसलिए, जीन समारोह के अध्ययन के लिए कई अन्य तकनीकों की तुलना में, आरएनएआई तेज, सस्ती है, और बड़े पैमाने पर स्क्रीन 23-25 के लिए आवेदन किया जा सकता है।
आरएनएआई की व्यापक उपयोगिता beyon अध्ययन के लिए उपलब्ध प्रजातियों की रेंज का विस्तार, जीवों की एक विस्तृत श्रृंखला में कार्यात्मक अध्ययन बाहर ले जाने के लिए साधन प्रदान करता हैपारंपरिक मॉडल प्रणाली है जिसके लिए आनुवंशिक उपकरण विकसित किया गया है घ। उदाहरण के लिए, गैर मॉडल प्रणाली का उपयोग अध्ययन विभिन्न विकास मोड का प्रतिनिधित्व करने या अलग रूपात्मक सुविधाओं 26-29 प्रदर्शन प्रजातियों से orthologs के कार्यों की तुलना द्वारा जीन और जीन नेटवर्क के विकास में अंतर्दृष्टि देने के लिए आवश्यक हैं। अध्ययन के इन प्रकार के जैविक विविधता का एक बेहतर समझ प्रदान करेगा, दोनों एप्लाइड और बुनियादी अनुसंधान के लिए प्रभावों के साथ।
ग्रह पर सबसे बड़ा पशु समूह होने के नाते, कीड़े अंतर्निहित तंत्र विविधता का पता लगाने के लिए एक महान अवसर प्रदान करते हैं। इसके अतिरिक्त, कीड़े आम तौर पर छोटे हैं, कम जीवन चक्र, उच्च उपजाऊपन है, और प्रयोगशाला में पीछे करने के लिए आसान कर रहे हैं। पिछले दो दशकों में, आरएनएआई सफलतापूर्वक आदेशों फैले, Diptera (सच मक्खियों) 5, Lepidoptera (तितलियों और कीटों), 30 सहित कीड़ों में लागू किया गया है, Coleoptera (बीटल) 16,31, कलापक्ष (sawfझूठ, ततैया, चींटियों और मधुमक्खियों) 32, Hemiptera (सच कीड़े), Isoptera (दीमक) 34, Blattodea (तिलचट्टे) 35, ऋजुपक्ष कीटवर्ग (क्रिकेट, टिड्डे, टिड्डियां, और katydids) 36 और Phthiraptera (जूँ) 37। आरएनएआई के सफल आवेदन जल्दी embryogenesis में patterning के अध्ययन के लिए कार्यात्मक डेटा (पूर्वकाल पीछे अक्ष 32, पृष्ठीय उदर अक्ष 28, विभाजन 26,38), लिंग निर्धारण 39,40, काइटिन / छल्ली जैवसंश्लेषण 41 प्रदान की गई है, ecdysone 42 सिगनल, सामाजिक व्यवहार 43, और अधिक। विभिन्न कीट प्रजातियों के लिए विकसित आरएनएआई तरीकों अतिरिक्त लाभ यह है कि वे कीट नियंत्रण (44-46 में समीक्षा) के लिए उपयोगी होने की संभावना है हो सकता है। आरएनएआई प्रभाव जीन विशिष्ट के साथ-साथ प्रजाति विशेष के लिए किया जाएगा, जब तक कि गैर संरक्षित क्षेत्रों को लक्षित करने के लिए चुना जाता है। मधुमक्खियों और कीड़ों की तरह लाभकारी कीट प्रजातियों, लक्षित करने के अस्तित्व के लिए महत्वपूर्ण जीनों के लिएवायरस या परजीवी संक्रमण को नियंत्रित करने के लिए इन प्रजातियों 47,48 रक्षा के लिए एक उपन्यास रणनीति प्रदान कर सकता है।
Dermestes maculatus (डी maculatus), आम का नाम छिपाने बीटल, अंटार्कटिका को छोड़कर दुनिया भर में वितरित किया जाता है। एक holometabolous कीट के रूप में, डी maculatus जीवन चक्र भ्रूण, लार्वा, पोटा संबंधी, और वयस्क चरणों (चित्रा 1) शामिल हैं। यह शरीर पर खिलाती है क्योंकि, डी maculatus मरे हुए जानवरों पांडुलेख करने के लिए संग्रहालयों में प्रयोग किया जाता है और फॉरेंसिक entomologists मौत 49,50 के समय का अनुमान लगाने के लिए इसका इस्तेमाल कर सकते हैं। डी maculatus शवों, सूखे मांस, पनीर, और pupae / अन्य कीड़ों के ककून सहित पशु उत्पादों पर खिलाती है और इस तरह के परिवारों को नुकसान, संग्रहीत भोजन, और रेशम, पनीर, मांस और उद्योगों 51,52 कारण बनता है। इस बीटल में आरएनएआई को लागू करने में अपनी आर्थिक प्रभाव को कम करने के लिए एक कुशल और पर्यावरण के अनुकूल तरीके प्रदान कर सकता है। हमारी प्रयोगशाला में एक नया मीटर के रूप में डी maculatus इस्तेमाल किया गया हैOdel कीट विभाजन 53 अध्ययन करने के लिए। प्रयोगशाला के पालन करने के लिए उत्तरदायी होने के अलावा, डी maculatus के रूप में यह एक मध्यवर्ती रोगाणु डेवलपर है, यह एक उपयोगी प्रजातियों बनाने छोटी और लंबी रोगाणु विकास के बीच संक्रमण का अध्ययन करने के लिए बुनियादी अनुसंधान के लिए ब्याज की है।
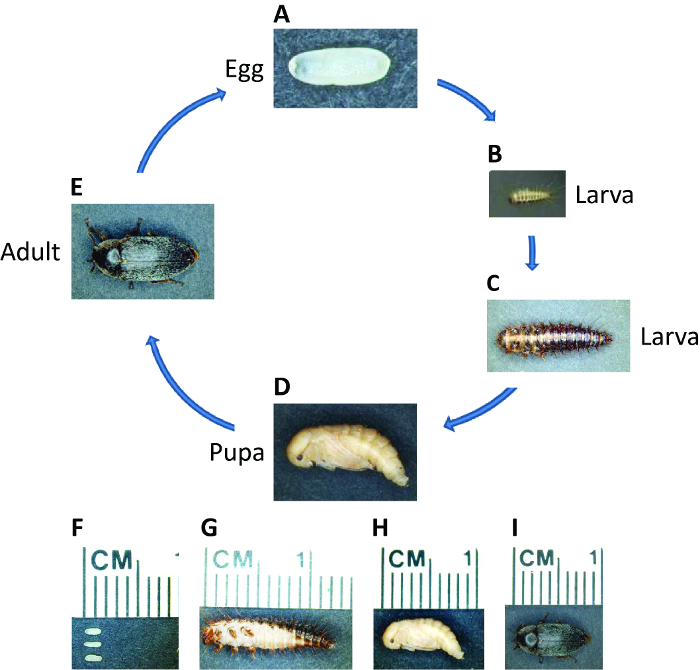
चित्रा 1: डी maculatus के जीवन चक्र। विभिन्न चरणों में जीवन डी maculatus के फोटोग्राफ, के रूप में संकेत दिया। वयस्क के लिए अंडे से जीवन चक्र कम तापमान पर 30 डिग्री सेल्सियस पर तीन सप्ताह लेकिन अब लगता है। (ए, एफ) हौसले रखी भ्रूण हल्के पीले रंग और अंडाकार, लंबाई में लगभग 1.5 मिमी के लिए सफेद होते हैं। Embryogenesis लेता है ~ 30 डिग्री सेल्सियस पर 55 घंटा। (बी, सी और जी) लार्वा अंधेरे pigmented धारियों और setae के साथ कवर किया जाता है। लार्वा पर्यावरण और उनकी लंबाई के आधार पर कई instars के माध्यम से जाने पर 1 सेमी तक विस्तार कर सकते हैं। (डी, एच) </strओंग> युवा pupae हल्के पीले रंग की हैं। 30 डिग्री सेल्सियस पर 7 दिन – pupation ~ 5 लेता है। (ई, मैं) eclosion के फौरन बाद, अंधेरे रंजकता वयस्क बीटल शरीर पर प्रकट होता है। वयस्क कई महीनों तक रह सकते हैं और एक महिला उसकी जीवन भर से अधिक भ्रूण के सैकड़ों रखना कर सकते हैं। यह आंकड़ा का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
इससे पहले, हम पता चला कि आरएनएआई डी maculatus 53 में जीन समारोह नीचे दस्तक करने में प्रभावी है। यहाँ हमारे अनुभव प्रयोगशाला में डी maculatus कालोनियों पालन दोनों भ्रूण और माता पिता का आरएनएआई सेट-अप, इंजेक्शन, बाद इंजेक्शन की देखभाल, और प्ररूपी विश्लेषण के लिए कदम-दर-कदम प्रोटोकॉल के साथ साझा किया जाता है। यहां पेश dsRNA की मध्यस्थता जीन पछाड़ना और विश्लेषण के तरीकों न केवल डी maculatus में प्रश्नों के समाधान के लिए विस्तृत जानकारी प्रदान करते हैं, लेकिन यह भी के लिए संभावित महत्व हैआर अन्य गैर-मॉडल बीटल / कीट प्रजातियों में आरएनएआई आवेदन।
