La feuille de maïs est un modèle idéal pour étudier la formation des champs de développement au cours de la morphogenèse, car il a une frontière distincte entre la lame et la gaine qui se prête à la dissection génétique (Figure 1A). Pendant les premiers stades de développement de la feuille, une bande linéaire de cellules plus petites, la bande de preligule (PLB), subdivise l'ébauche de feuille en pré-lame et pré-gaine domaines. Une ligule frange semblable et oreillettes triangulaires se développent à partir de la PLB (Figure 1A, C, D). écrans génétiques ont identifié des mutations qui perturbent la limite lame-gaine. Par exemple, liguleless1 récessive (LG1) mutations supprimer la ligule et oreillettes 1, 2, 3, 4 (figure 1B). L'hybridation in situ a révélé que la transcription lg1 accumule au PLB et ligule émergents, ce qui en fait un excellent marqueur pour le développement ligule 5, 6 (figure 1E).
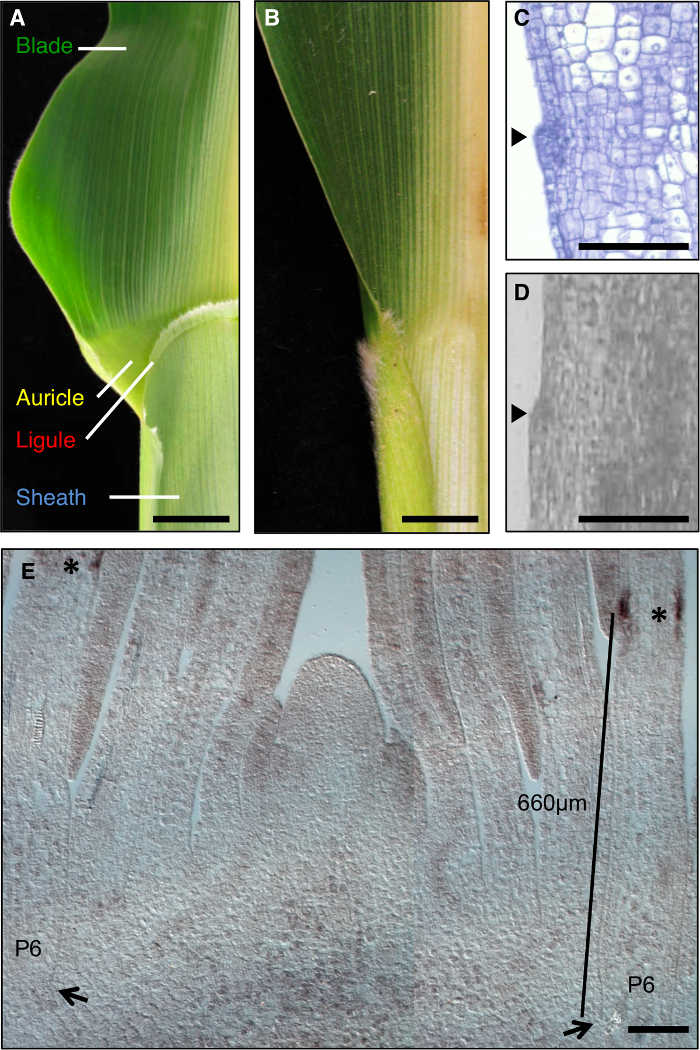
Figure 1: type sauvage et liguleless1-R feuilles de maïs. (A) région limite Blade-gaine de maturité des feuilles de type sauvage montrant les structures de ligule et oreillette. (B) de la région de la frontière Blade-gaine mature liguleless1-R feuille montrant l' absence de structures de ligule et oreillette. Feuilles en A et B ont été coupés en deux le long de la nervure médiane. (C) longitudinale coupe de type sauvage feuille primordium. L'échantillon a été traité et colorées pour l'analyse histologique. Initier la ligule ressort comme une bosse en saillie par rapport au plan de la lame (pointe de flèche). (D) sect longitudinaleion par type sauvage feuille primordium. L'échantillon a été traité pour LM, comme décrit dans le texte. Arrowhead indique lancer ligule. (E) LG1 hybridation in situ de la tige latérale en coupe longitudinale de sommet en. Les astérisques indiquent l' accumulation de transcription lg1 au PLB du primordium P6 feuille. Les flèches indiquent la base de P6 primordium. Bar indique la mesure de la base de l'ébauche à la PLB. Les barres d'échelle en A et B = 20 mm. Les barres d'échelle dans CE = 100 um. Ce chiffre a été modifié depuis la référence 6 (Droit d' auteur Société américaine des biologistes végétaux). S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
Dans cette étude, LM ARN-Seq a été utilisée pour identifier un ensemble de gènes qui sont exprimés de manière différentielle (DE) à la limite lame-gaine par rapport à d'autres parties du primordium foliaire et Ide gènes ntifier qui sont dans des mutants DE lg1-R par rapport aux frères et sœurs de type sauvage. LM ARN-Seq est une méthode de quantification de l' accumulation du transcrit dans des cellules spécifiques ou des domaines cellulaires 7. Les systèmes LM combinent un laser et un microscope avec un appareil photo numérique. tissus Sectionné est monté sur des lames et vu à travers le microscope. Le logiciel de LM comprend généralement des outils de dessin qui permettent à l'utilisateur d'esquisser une région sélectionnée pour microdissection. Les coupes laser le long de la ligne, et le tissu sélectionné est catapulté hors de la diapositive et dans un tube suspendu au-dessus de la diapositive. LM permet à l'utilisateur de microdissect domaines précis, y compris les couches de cellules spécifiques et même des cellules individuelles 8, 9. L'ARN peut alors être extrait du tissu microdissection. Par la suite, le composant ARN-Seq utilise le séquençage de la prochaine génération à la séquence des banques d' ADNc générées à partir de l'ARN extrait 10,= "xref"> 11.
Les principaux avantages de LM ARN-seq sont la capacité à quantifier l' accumulation des transcrits dans des domaines bien définis et la capacité de profiler l'ensemble transcriptome simultanément 7. La technique est particulièrement adaptée à sonder les événements précoces du développement, où la région d'intérêt est souvent microscopique. Des études antérieures ont utilisé LM combiné avec la technologie des biopuces pour étudier les processus de développement dans les plantes 9, 12, 13. ARN-Seq a l'avantage de quantifier les transcriptions à travers une large gamme dynamique, y compris les gènes de faible exprimés, et de l' information de séquence préalable est pas nécessaire 10, 11. En outre, LM ARN-Seq a le potentiel de mettre en évidence les gènes importants pour le développement qui peuvent manquer dans les écrans de mutagenèse en raison de la redondance génétique ou à la létalité de la perte-ofmutant de fonction.
Gènes importants pour le développement, tels que sheath1 étroite (NS1) et en forme de cuvette de cotyledon2 (CUC2), ont souvent des profils d'expression spécifiques d'un seul ou de quelques cellules 17, 18, 19, 20. Beaucoup sont exprimés seulement au cours des étapes précoces du développement et non pas dans l'organe mature. Lorsque des organes entiers ou de grands domaines sont analysés, ces transcrits spécifiques des cellules sont diluées et ne peuvent pas être détectées dans les analyses plus classiques. En permettant des analyses des domaines bien définis, LM ARN-Seq permet à ces gènes spécifiques de tissus à être identifiés et quantifiés.
Les facteurs cruciaux dans le succès des expériences décrites ici ont une analyse histologique approfondie qui a guidé la sélection de l'étape de développement approprié et le domaine de l'analyse, et MeasureMe précisent de domaines de cellules tissulaires pour LM. Pour veiller à ce que les domaines équivalents ont été échantillonnés pour toutes les répétitions, les tissus ont été recueillis à partir de primordiums foliaires au même stade de développement et les domaines microdissection ont été mesurés par rapport aux repères morphologiques telles que la ligule émergente (figure 2). Il est connu que certains gènes sont exprimés dans un gradient allant de la pointe à la base de la feuille. En mesurant les domaines précis, la variation due à l' échantillonnage à différents endroits le long de l'axe proximal-distal feuille a été maintenu à un minimum (figure 3A). Par microdissecting domaines de la même taille, la variation due à la différence de dilution des produits de transcription spécifiques de cellules a également été réduite (figure 3B). des sections longitudinales latérales du sommet de pousses ont été utilisées pour toutes microdissections. Ceux – ci sont des profilés qui sont perpendiculaires à l'axe de la nervure médiane marge (figure 4). En utilisant uniquement les sections qui incluent le SAM assure que les régions latérales équivalentes deprimordia foliaires sont analysés.
Dans les échantillons traités et sectionnés pour LM, le premier signe morphologique d'excroissance ligule est une bosse sur le côté adaxial en raison de divisions cellulaires périclines dans l'épiderme adaxiales (figure 1D, Figure 2). Il a été déterminé que la ligule émergente pourrait être identifié de manière fiable à plastochron 7 primordia foliaires de scène. Nous étions intéressés par les gènes exprimés dans toute la région, y compris le ligule ligule émergents et les cellules immédiatement distales qui formeront l'auricule. Afin de s'assurer que les pièces de tissus équivalents ont été faites, la bosse ligule a été utilisé comme point de repère morphologique et un rectangle de 100 um centrée sur la bosse ligule a été sélectionné pour LM (figure 2A, 2B). rectangles de taille équivalentes de pré-lame et pré-gaine ont été choisis parmi les mêmes primordia foliaires.
Les analyses des plantes mutantes ont présenté une liguleless challe différentnge; mutants lg1-R ne forment pas une ligule, donc cette fonctionnalité morphologique ne peut pas être utilisé pour sélectionner la région pour LM. Au lieu de cela, le domaine de l' accumulation de transcription lg1 dans de type sauvage primordiums foliaires a été déterminé, et une région qui engloberait ce domaine a été défini. Ces analyses préliminaires ont été effectuées sur des plants de la même plantation qui ont été utilisés pour l'analyse finale, puisque les travaux antérieurs ont montré que l'emplacement de la PLB varie en fonction des conditions de croissance. L'hybridation in situ a indiqué que les transcriptions LG1 accumulent dans la PLB de P6 primordiums foliaires (figure 1E). Nous avons choisi un domaine 400-900 um à partir de la base de la primordia feuille qui englobait le domaine d'expression lg1 (rectangles violets, figure 2A) et capturé ces régions équivalentes de type sauvage et les plantes lg1-R. Pour réduire au minimum la variation des conditions de fond et de croissance génétique lorsque l'on compare transcriptaset accumulation dans lg1-R et des plantes de type sauvage, ségrégation des familles de mutants et frères et sœurs de type sauvage ont été utilisés.
