A folha de milho é um modelo ideal para estudar a formação de campos de desenvolvimento durante a morfogénese, uma vez que tem uma fronteira distinta entre a lâmina e a bainha que é passível de dissecção genética (Figura 1A). Durante as fases iniciais do desenvolvimento foliar, uma faixa linear de células menores, a banda preligule (PLB), subdivide o primórdio foliar em domínios pré-lâmina e pré-bainha. A ligule franja-like e aurículas triangulares desenvolver a partir da PLB (Figura 1A, C, D). telas genéticas identificaram mutações que perturbam o limite lâmina de bainha. Por exemplo, liguleless1 recessivo (LG1) mutações excluir o ligule e aurículas 1, 2, 3, 4 (Figura 1B). A hibridação in situ revelou que LG1 transcrição acumula no PLB e ligule emergentes, tornando-se um excelente marcador para o desenvolvimento ligule 5, 6 (Figura 1E).
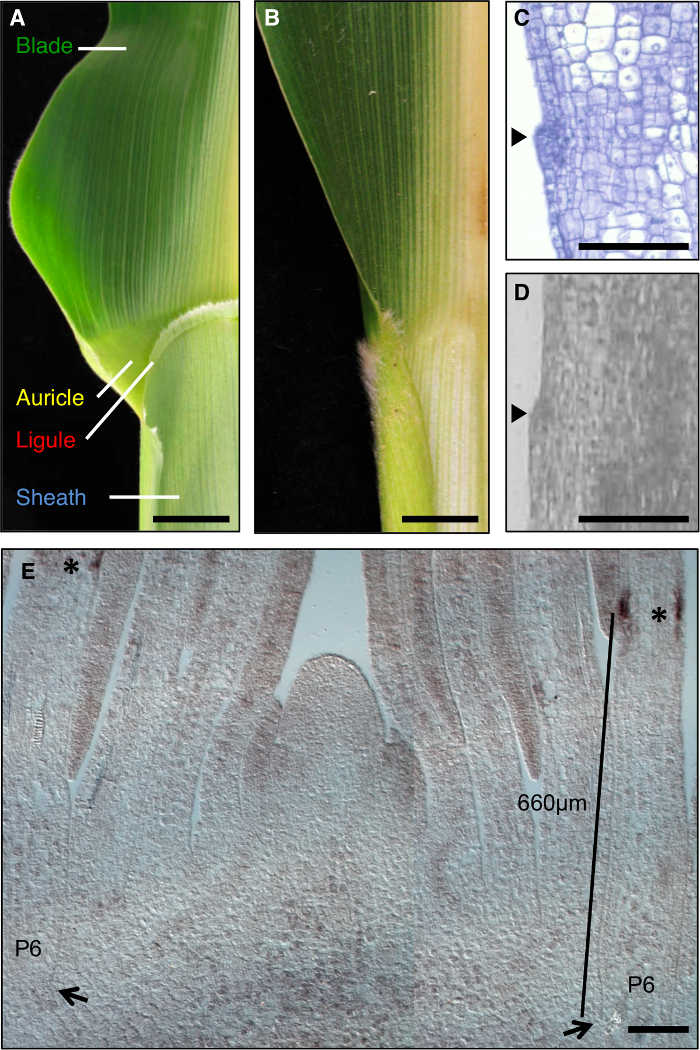
Figura 1: Tipo selvagem e folhas de milho liguleless1-R. (A) a região Blade-bainha limite da folha de tipo selvagem maduro mostrando estruturas ligule e aurícula. (B) a região Blade-bainha limite da maturidade liguleless1-R folha que mostra ausência de estruturas ligule e aurícula. Folhas em A e B foram cortadas pela metade ao longo da nervura central. (C) longitudinal secção através primórdio foliar de tipo selvagem. Amostra foi processada e corados para análise histológica. A lígula iniciadora é evidente como uma colisão que sobressai a partir do plano da folha (ponta de seta). (D) seita Longitudinalíon através primórdio folha de tipo selvagem. Amostra foi processada por LM como descrito no texto. Arrowhead indica iniciar ligule. (E) lg1 hibridização in situ de corte longitudinal lateral do ápice filmagem. Os asteriscos indicam acumulação transcrição lg1 na PLB do primórdio foliar P6. As setas indicam a base do P6 primórdio. A barra indica a medição a partir da base de o primórdio da PLB. As barras de escala em A e B = 20 mm. As barras de escala em Ce = 100 uM. Esta figura foi modificado a partir da referência 6 (Copyright: Sociedade Americana de biólogos da planta). Por favor clique aqui para ver uma versão maior desta figura.
Neste estudo, LM RNA-Seq foi empregue para identificar um conjunto de genes que são expressos diferencialmente (DE) para a lâmina de bainha limite em relação a outras partes do primórdio foliar e de IDE genes ntify que estão DE em mutantes lg1-R em relação aos irmãos de tipo selvagem. LM RNA-Seq é um método de quantificação de acumulação de transcrição em células específicas ou domínios celulares 7. sistemas lineares combinar um laser e um microscópio com uma câmera digital. tecido é seccionado montadas em lâminas e visualizaram pelo microscópio. O software LM normalmente inclui ferramentas de desenho que permitem ao usuário para delinear qualquer região selecionada para microdissection. Os cortes a laser ao longo da linha, e o tecido selecionado é catapultado para fora da lâmina e em um tubo suspenso acima do slide. LM permite que o usuário microdissect domínios precisos, incluindo camadas de células específicas e até mesmo células individuais 8, 9. ARN pode então ser extraído a partir do tecido microdissecados. Subsequentemente, o componente de RNA-Seq utiliza a próxima geração de sequenciação para sequenciar bibliotecas de cDNA geradas a partir do RNA extraído 10,= "xref"> 11.
Principais vantagens do LM RNA-seq são a capacidade de quantificar a acumulação de transcrição em domínios bem definidos ea capacidade ao perfil todo o transcriptoma simultaneamente 7. A técnica é particularmente adequada para sondagem eventos precoces de desenvolvimento, onde a região de interesse é muitas vezes microscópica. Estudos anteriores têm utilizado LM combinado com a tecnologia de microarrays para estudar processos de desenvolvimento em plantas 9, 12, 13. RNA-Seq tem a vantagem de quantificar transcritos de uma vasta gama dinâmica, incluindo os genes de baixo-expresso, e informação da sequência anterior não é necessária 10, 11. Além disso, LM RNA-Seq tem o potencial para destacar genes desenvolvente importantes que podem ser perdidas em telas de mutagénese devido à redundância genética ou a letalidade da perda de de-função mutante.
Genes Developmentally importantes, como sheath1 estreita (NS1) e em forma de taça cotyledon2 (CUC2), muitas vezes têm padrões de expressão específicas de apenas uma ou algumas células 17, 18, 19, 20. Muitos são expressos apenas durante as fases iniciais de desenvolvimento e não no órgão maduro. Quando órgãos inteiros ou grandes domínios são analisados, estes transcritos específicos de células são diluídas e não pode ser detectada em análises mais convencionais. Ao permitir análises de domínios bem definidos, LM RNA-Seq permite que estes genes específicos de tecido a ser identificados e quantificados.
factores cruciais para o sucesso das experiências descritas aqui foram uma análise histológica minuciosa que orientou a seleção do estágio de desenvolvimento apropriado e de domínio para análise e MeasureMe precisant de domínios de tecido celular para LM. Para assegurar que os domínios equivalentes foram amostrados de todas as repetições, o tecido foi recolhido a partir de primórdios foliares, ao mesmo estágio de desenvolvimento e os domínios microdissecadas foram medidos em relação a pontos de referência morfológicas, tais como a lígula emergente (Figura 2). Sabe-se que alguns genes são expressos em um gradiente a partir da ponta para a base da folha. Ao medir domínios precisos, devido à variação de amostragem a partir de diferentes locais ao longo do eixo proximal-distal folha foi mantido a um mínimo (Figura 3A). Por microdissecting domínios do mesmo tamanho, devido a variação diferencial de diluição de transcritos específicos de células também foi reduzida (Figura 3B). secções laterais longitudinais do ápice filmagem foram usadas para todos os microdissecações. Estes são secções que são perpendiculares ao eixo nervura central-margem (Figura 4). Usando apenas as seções que incluem a SAM assegura que as regiões laterais equivalentes deprimórdios foliares são analisados.
Em amostras processadas e seccionados para LM, o primeiro sinal morfológico de crescimento ligule é um inchaço no lado adaxial devido a divisões celulares periclinais na epiderme adaxial (Figura 1D, Figura 2). Determinou-se que o ligule emergentes poderiam ser identificados de forma confiável no plastocrono 7 primórdios foliares palco. Nós estávamos interessados em genes expressos em toda a região ligule, incluindo a ligule emergentes e as células imediatamente distais que formarão a aurícula. A fim de assegurar que as selecções de tecido equivalente foram feitas, a colisão lígula foi usada como um ponto de referência morfológica e um rectângulo de 100 um centrada na protuberância lígula foi seleccionado para LM (Figura 2A, 2B). rectângulos de dimensão equivalente de pré-lâmina e pré-bainha foram seleccionados a partir dos mesmos primórdios foliares.
As análises de plantas mutantes liguleless apresentada uma diferente challeESL; lg1-R mutantes não formam um ligule, portanto, esse recurso morfológica não poderia ser usado para selecionar a região de LM. Em vez disso, foi determinado o domínio de transcrição acumulação LG1 em primórdios foliares de tipo selvagem, e uma região que abrange este domínio foi definido. Estas análises preliminares foram realizadas em plântulas a partir do mesmo plantação como foram usadas para a análise final, uma vez que o trabalho anterior demonstrou que a localização do PLB varia dependendo das condições de crescimento. A hibridação in situ indicaram que os transcritos LG1 acumulam no PLB de P6 primórdios foliares (Figura 1E). Nós seleccionado um domínio de 400-900 ^ M a partir da base do primórdios foliares, que englobava o domínio de expressão LG1 (rectângulos roxo, Figura 2A) e capturou estas regiões equivalentes do de tipo selvagem e plantas LG1-R. Para minimizar a variação de fundo e de crescimento genéticos condições quando se comparam transcripforam usadas acumulação t em LG1-R e plantas de tipo selvagem, segregando famílias de mutantes e de tipo selvagem irmãos.
