Лист кукурузы является идеальной моделью для исследования образования полей развития в процессе морфогенеза, поскольку она имеет четкую границу между лопаткой и оболочкой, которая поддается генетической диссекции (рис 1А). На ранних стадиях развития листьев, линейной полосы более мелких ячеек, то preligule группа (PLB), подразделяет листовой зачаток в предварительно лопаточной и предварительно оболочки доменов. Бахромой, как Язычок и треугольные ушки развиваются из PLB (рис 1А, C, D). Генетические экраны выявили мутации, которые нарушают границу лопастного оболочки. Например, рецессивный liguleless1 (lg1) мутации удаления язычка и ушные раковины 1, 2, 3, 4 (Фигура 1В). В гибридизация показало , что lg1 транскрипт накапливается в РLB и возникающих язычка, что делает его отличным маркером для язычка развития 5, 6 (рис 1E).
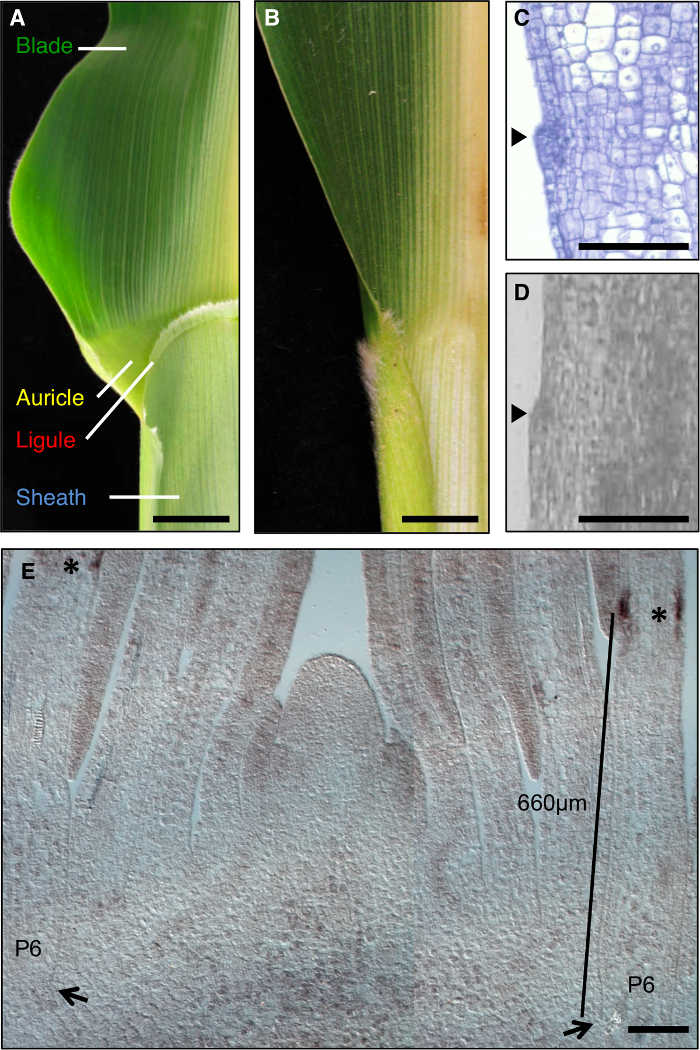
Рисунок 1: дикого типа и liguleless1-R листьев кукурузы. (А) граница Блейд-оболочка область зрелого дикого типа листа , показывающий язычка и предсердия структур. (B) граница Блейд-оболочка область зрелого liguleless1-R листьев показывая отсутствие язычка и ушной раковины структур. Листья в А и В были сокращены в два раза вдоль жилки. (С) Продольный разрез через лист примордия дикого типа. Образец был обработан и окрашивают для гистологического анализа. Инициирующий Язычок очевидно как выпуклость, выступающую из плоскости листа (стрелки). (D) Продольная разделиона через лист зачатка дикого типа. Образец был обработан для LM, как описано в тексте. Arrowhead указывает на инициирование язычка. (E) lg1 на месте гибридизации побегов верхушки корня бокового продольного разреза в. Звездочки показывают lg1 накопление транскриптов в PLB от P6 листа зачатка. Стрелки указывают основание P6 зачатка. Бар показывает измерение от основания зачатка к ИПР. Шкала баров в А и В = 20 мм. Шкала баров в CE = 100 мкм. Эта цифра была изменена из ссылки 6 (Авторское право Американского общества биологов растений). Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
В этом исследовании, Л.М. Секвенирование РНК был использован, чтобы идентифицировать набор генов, которые дифференциально экспрессируются (DE) на границе лопастного оболочки по отношению к другим частям листа зачатка и Ide ntify гены, которые DE в lg1-R мутантов по отношению к братьев и сестер дикого типа. LM-РНК Seq представляет собой метод количественного накопления транскриптов в специфических клетках или клеточных доменов 7. LM системы сочетают в себе лазер и микроскоп с цифровой камерой. Секционного ткань монтируется на салазках и просмотрены через микроскоп. Программное обеспечение LM обычно включает в себя инструменты для рисования, которые позволяют пользователю выделить любую выбранную область для микродиссекции. Разрезы лазерные вдоль линии, и выбранная ткань катапультировался за пределы слайда и в трубу, подвешенную над слайде. LM позволяет пользователю microdissect точные домены, в том числе специфических клеточных слоев и даже отдельных клеток 8, 9. РНК затем может быть извлечено из микродиссекции ткани. Затем РНК-Seq компонент использует следующего поколения последовательности в последовательности кДНК – библиотек , сгенерированные из извлеченной РНК 10,= "Xref"> 11.
Основные преимущества LM Секвенирование РНК являются способность количественного накопления транскриптов в четко определенных областях и способность к профилю весь транскриптом одновременно 7. Метод особенно подходит для зондирования ранних событий в области развития, где область интереса часто микроскопическими. Предыдущие исследования использовали LM в сочетании с микрочипов технологии для изучения процессов развития растений 9, 12, 13. РНК-Seq имеет преимущество количественной оценки транскриптов в широком динамическом диапазоне, в том числе с низкой экспрессируемых генов, и информация до последовательности не требуется 10, 11. Более того, LM-Seq РНК имеет потенциал, чтобы выделить важные гены уровню развития, которые могут быть пропущены в экранах мутагенеза из-за генетической избыточности или к летальности потерь посещающихфункция мутант.
Развивающих важные гены, такие как узкий sheath1 (ns1) и чашеобразной cotyledon2 (cuc2), часто имеют специфические паттерны экспрессии только один или несколько ячеек 17, 18, 19, 20. Многие выражаются только на ранних стадиях развития, а не в зрелом органе. Когда целые органы или крупные домены анализируются, эти клеточные специфические транскрипты разбавлены и не могут быть обнаружены в более обычных анализах. Допустив анализ точно определенных областей, LM-Seq РНК позволяет эти гены тканеспецифические быть идентифицированы и количественно.
Решающие факторы в успехе экспериментов, описанных здесь, были тщательный гистологический анализ, который руководствовался выбор соответствующей стадии развития и области для анализа и точного MeasureMeнт доменов клеточной ткани для LM. Для того, чтобы гарантировать , что эквивалентные домены были отобраны для всех повторах, ткань была собрана из листьев зачатков на той же стадии развития и микродиссекции домены были измерены относительно морфологических ориентиров , таких как формирующейся язычка (рис 2). Известно, что некоторые гены экспрессируются в градиенте от кончика к основанию листа. Измеряя точные домены, изменение из – за выборки из разных мест вдоль листа проксимальной-дистальной оси была сведена к минимальной (рис 3А). По microdissecting домены одного и того же размера, изменение за счет дифференциальной разбавление клеток специфических транскриптов также была снижена (фигура 3В). Боковые продольные участки верхушки побега были использованы для всех microdissections. Эти участки, перпендикулярные оси жилка рентабельностью (Рисунок 4). Используя только разделы, которые включают SAM гарантирует, что эквивалентные боковые участкилистовые зачатки анализируются.
В образцах , обработанных и секционного для LM, первым морфологическим признаком язычка выроста является шишка на адаксиальную стороне из – за периклинальных клеточных делений в адаксиальной эпидермиса (рис 1D, Рисунок 2). Было установлено, что формирующаяся Язычок может быть надежно идентифицирован на plastochron 7 этап листьев зачатков. Мы были заинтересованы в генах, выраженных во всем регионе язычка, включая формирующейся язычка и клетки немедленно дистальных, которые будут образовывать ушную раковину. Для того , чтобы гарантировать , что выборы эквивалентные ткани были сделаны, Язычок шишка был использован в качестве ориентира морфологического и прямоугольник 100 мкм с серединой на Язычок бугорок был выбран для LM (рис 2А, 2В). Эквивалент размера прямоугольниками предварительного лезвия и предварительной оболочки были выбраны из тех же листьев зачатков.
Анализ liguleless мутантных растений представлены различные ChalleНге; lg1-R мутанты не образуют язычка, поэтому эта морфологическая особенность не может быть использована для выбора региона для LM. Вместо этого область lg1 накопления транскриптов в листьев зачатков дикого типа была определена, и была определена область , которая охватывала бы эту область. Эти предварительные анализы проводили на проростках из того же посадки, которые были использованы для окончательного анализа, поскольку предыдущие исследования показали, что местоположение PLB изменяется в зависимости от условий роста. В гибридизация показали , что lg1 транскрипты накапливаются в PLB Р6 листьев зачатков (рис 1E). Мы выбрали домен 400-900 мкм от основания листа зачатков , который охватывает область lg1 выражения (фиолетовый прямоугольник, рисунок 2А) и захватил эти эквивалентные области из дикого типа и lg1-R растений. Чтобы свести к минимуму изменение генетического фона и условий роста при сравнении transcripбыли использованы т накопление в lg1-R и растений дикого типа, семейства разделения мутантов , братьев и сестер дикого типа.
