The following procedures adhere to a protocol that was approved by the Institutional Review Board of University of California, Davis; consult with the local ethics board prior to beginning research.
1. Isolate Primary Fibroblasts from Human ACL Remnants
NOTE: Maintain sterility and perform all steps in a biological safety cabinet (BSC).
- Obtain approval from the appropriate ethics review board for the collection and use of human tissues as described below.
- Prepare 5x antibiotic/antimycotic (ABAM) solution by diluting 100x antibiotic/antimycotic solution in 1X phosphate-buffered saline (PBS)
- Collect ACL tissue fragments in 5X ABAM solution in a 50 mL conical tube, store at 4 °C until the digestion step. Cut tissue into smaller fragments with a razor blade if necessary to a maximum size of 1 x 1 x 1 cm3.
CAUTION: Comply with local biohazard regulations for proper use of biohazard material and decontamination and disposal of biohazard waste. - Prepare a sufficient volume of collagenase solution to submerge the tissue fragments. Dissolve collagenase type II (1 mg/mL) in high glucose Dulbecco's modified Eagle medium (DMEM) containing 1x penicillin/streptomycin, and 20% fetal bovine serum (FBS) and filter at 0.22 µm.
- Rinse ACL tissue 3 times in PBS.
- Digest tissue. Transfer ACL tissue to a new 50 mL conical tube and add a sufficient volume of collagenase solution to submerge the tissue. Incubate at 37 °C overnight (~17 h).
- Before the digestion duration is complete, prepare growth media (GM) by supplementing DMEM high glucose media with 10% FBS and 100 U/mL penicillin.
- 15 min before the digestion time is complete, briefly vortex the tissue-containing 50 mL tube 3 times every 5 min.
- Using a 25 mL serological pipette, triturate the digested tissue vigorously to break the tissue up further and dislodge cells.
- Centrifuge at 1,500 x g for 5 min. Aspirate the supernatant and resuspend pellet in 10 mL GM.
- Repeat steps 1.8-1.9 three more times.
- After the last centrifugation, resuspend the pellet in 5-10 mL GM. Use a small sample of the cell suspension to perform a cell count with a hemocytometer and assess cell viability using Trypan Blue.
- Plate the cell suspension onto 15 cm tissue culture plates at a density of 3-4 x 105 cells per plate.
- Culture to 70% confluence in a sterile incubator maintained at 37 °C and 5% CO2, changing the GM every three days. Use or store (see below) cells within 5 passages.
- Freeze cells for future use.
- Trypsinize cells at 70% confluence as follows. Aspirate media and wash cells with PBS. Add enough pre-warmed (37 °C) 0.05% trypsin to just cover the bottom of tissue culture plates. Place plates in the culture incubator for ~5 min until cells are detached (verify cells are floating using a light microscope). Use a pipette to collect cells and dispense into a Falcon tube.
- Centrifuge to pellet cells, and resuspend cells in DMEM high glucose media containing 20% FBS and 10% dimethyl sulfoxide (DMSO). Aliquot cell suspension into cryovials and cool at -1 °C/min for at least 24 h. Store frozen cryovials in liquid nitrogen.
2. Prepare Silicone-coated Plates
- Prepare 35 mm tissue culture plates: Remove lids and lay out open plates on a flat surface.
- Mix silicone elastomer kit according to manufacturer's instructions.
- Use a 10 mL syringe to dispense approximately 2 mL per 35 mm plate.
- Allow silicone to cure at room temperature for 2-3 days.
3. Prepare Brushite Cement Anchors
- In advance, prepare silicone reverse-molds containing cylindrical wells for anchor formation. Reverse molds can be made to the specifications of the desired anchor shape and size.
- Determine the desired height and diameter of the final anchor. This protocol uses a custom-made mold formed from craft silicone in a 35 mm tissue culture dish in which plastic cylinders of about 3.25 mm in diameter were placed, allowing for a final mold height of about 6.5 mm. Final anchor dimensions are approximately 3-3.5 mm in height and about 3.4 mm in diameter with 1.5 mm pin protruding from the bottom of the anchor.
- Add uncured silicone to a 35 mm dish in which the mold will be made. Place plastic cylinders accordingly.
NOTE: The size of the plastic cylinders will determine the diameter of the final anchors. The placement of the plastic cylinders and the amount of silicone used can be modified to produce anchors of different heights. The thickness between the bottom of the wells in the mold and the bottom of the mold itself will determine how much of the pin can protrude from the bottom of the anchor allowing the anchor to later be pinned securely in the silicone-coated dish. - After allowing the silicone to cure, remove the plastic cylinders and remove the mold from the 35 mm dish.
- Prepare a 3.5 M orthophosphoric/100 mM citric acid solution. Dissolve 0.961 g citric acid in 11.5 mL orthophosphoric acid. Bring the volume of the solution up to 50 mL with MilliQ water. Store solution at room temperature and protect from light.
- Prepare molds: Place one minutien pin in the center of each cylindrical well in the molds.
- Combine β-tricalcium phosphate and orthophosphoric/citric acid solution at a 1 g/mL concentration in a plastic weigh boat on ice.
- Mix the cement vigorously using a plastic cell scraper.
- Triturate the cement to continue mixing and pipette mixture into the mold
- Centrifuge the filled mold for 1 min at 2,250 x g.
- Allow brushite cement anchors to set at room temperature overnight.
- Remove anchors from the mold and pin two anchors 12 mm apart in each silicone-coated plate.
- Sterilize pinned plates by spraying with 70% ethanol, filling both the plates and lids, and placing into a BSC. After at least 30 min, aspirate plates and replace lids, storing in the BSC until needed.
4. Obtain Human Serum
- Ensure that approval by the appropriate ethics review board has been obtained for this protocol.
- Ensure that written informed consent has been obtained from human subjects to participate in a given intervention (e.g., exercise, food or drug intervention) that will effect desired changes in serum. Here, we describe the collection at rest and 15 min after resistance exercise.
- Using a trained phlebotomist, obtain a resting blood sample from a participant by venipuncture into an appropriate evacuated container.
- Collect a post-exercise blood sample 15 min after having the participants engage in the desired exercise protocol. As previously described1, use the resistance exercise protocol in this experiment to stimulate an endogenous biochemical response.
- Have participants perform five sets of leg press with one-minute rest between sets. Next, have the participants perform a set of knee extensions and a set of hamstring curls consecutively with no rest and then repeat the back-to-back exercises three times with 1 min rest between sets.
- Allow blood to clot before centrifuging at 1,500 x g for 10 min. Under sterile conditions, transfer serum to sterile tubes for future media supplementation (serum stored at 4 °C) and biochemical analysis (a small aliquot of serum stored at -20 °C).
5. Form Engineered Ligaments
NOTE: In advance, expand primary fibroblasts and prepare silicone-coated plates with pinned brushite anchors.
- Prepare reagents:
- Prepare thrombin. Dissolve bovine thrombin at 200 U/mL in DMEM high glucose media. Filter at 0.22 µm, aliquot, and store at -20 °C.
- Prepare fibrinogen. Dissolve bovine fibrinogen at 20 mg/mL in DMEM high glucose media. Incubate for 3-4 h in a 37 °C water bath, swirling every 30 min to aid dissolution. Filter at 0.22 µm (multiple filters may be needed), aliquot, and store at -20 °C.
- Prepare aprotinin. Dissolve aprotinin in 10 mg/mL in water. Filter at 0.22 µm, aliquot, and store at -20 °C.
- Prepare aminohexanoic acid. Dissolve aminohexanoic acid at 0.1g/mL in water. Filter at 0.22 µm, aliquot, and store at 4 °C.
- Prepare ascorbic acid. Dissolve ascorbic acid in DMEM high glucose media at a concentration of 50 mM. Filter at 0.22 µm and store at 4 °C.
- Prepare L-proline. Dissolve L-proline in PBS at a concentration of 50 mM. Filter at 0.22 µm and store at 4 °C.
- Prepare transformation growth factor-β1 (TGF-β1). Reconstitute TGF-β1 according to the manufacturer's directions at a concentration of 10 µg/mL. Aliquot and store at -20 °C.
- Determine the number of constructs required for the experiment and ensure sufficient numbers of pinned plates are prepared. Both biological and technical replicates are recommended. In the study1 highlighted here, use duplicate technical replicates and 12 biological replicates (serum from 12 individuals at rest and after exercise).
NOTE: Perform the following steps under sterile conditions in a BSC. - Expand cells by culturing in 15 cm plates to 70% confluence. 2.5 x 105 cells are required per construct.
- Trypsinize cells and resuspend in GM at a concentration of 3.67 x 105 cells/mL.
- Generate a master mix for the number of constructs required: For 1 construct, the master mix contains 681 µL cell suspension (containing 2.5 x 105 cells), 29 µL thrombin, 2 µL aprotinin, and 2 µL aminohexanoic acid.
- After mixing the master mix well, add 714 µL to each pinned plate in a 'figure 8' pattern around the brushite cement anchors. Ensure that the master mix directly contacts the sides of the anchors.
- Gently tap each plate to distribute the master mix evenly across the plate.
- For one plate at a time, quickly add 286 µL fibrinogen in a dropwise fashion evenly over one plate and immediately slide the plate back and forth and side to side over the surface of the BSC to distribute the fibrinogen to form the cell-embedded fibrin gels. Proceed to the next plate.
- Place the constructs in a sterile incubator maintained at 37 °C and 5% CO2 and incubate for at least 15 min to allow polymerization of the fibrinogen.
- Prepare sufficient feed media (FM) for 2 mL per construct. Supplement GM with 200 µM ascorbic acid, 50 µM proline, and 5 ng/mL TGF-β1.
- Add 2 mL FM to cover each construct. Culture the constructs in a sterile incubator maintained at 37 °C and 5% CO2 for a total of 14 days or to the desired endpoint, refreshing the media every second day with 2 mL FM
6. Tensile Testing Engineered Ligaments
NOTE: Tensile testing was performed using a custom-built tensile tester in a PBS bath; reverse-molded grips that are coupled to the force transducer hold brushite cement anchors in place during the test.
- Determine the length and width of the ligament constructs using digital calipers; calculate the cross-sectional area of the tissue.
- Unpin the ligament construct from the plate and place the anchors in the reverse molded grips, ensuring the construct is submerged in PBS.
- Adjust the distance between the grips, setting the length of the construct to its initial length.
- Begin the test: strain the construct to failure at a strain rate of 0.4 mm/s (or ~3%/s).
- After completion of the test, process the tissue remnants for collagen content (see section 7).
- From the resultant load-deformation data, calculate stress-strain data and quantify mechanical properties of interest; for example, maximal tensile load, ultimate tensile strength and modulus (i.e., elastic property over a linear region of the stress-strain curve).
7. Quantification of Collagen Content of Engineered Ligaments
- Remove engineered ligaments from brushite cement anchors and dry at 120 °C for 25 min.
- Determine the dry mass of constructs and place into individual 1.5 mL tubes. Dry constructs may be stored at room temperature until further processing.
- To each construct, add 200 µL 6 M HCl. Boil at 120 °C in a heating block for 2 h in a fume hood.
CAUTION: HCl is highly corrosive and acidic, the use of boil-proof/safe-lock tubes or other method of securing tubes is recommended. - Centrifuge the tubes briefly to collect liquid, and leave them uncapped to evaporate at 120 °C in a heating block for 1.5 h in a fume hood.
- Resuspend the resultant pellet in 200 µL hydroxyproline buffer. Store at -20 °C until needed.
- Prepare hydroxyproline buffer. In 300 mL water, add 16.6 g citric acid, 4 mL acetic acid, 11.4 g NaOH and stir until dissolved. pH to 6-6.5 and bring volume up to 500 mL. Add 250 µL toluene as a preservative and store at 4 °C protected from light.
- Prepare reagents.
- Prepare trans-4-Hydroxy-L-proline. Dissolve in water to make a 4 mg/mL solution.
- Prepare chloramine-T. Dissolve in water to make a 14.1 mg/mL solution.
- Prepare aldehyde-perchlorate. Dissolve 1.5 g 4-dimethylaminobenzaldehyde in 6 mL 1-propanol, 2.6 mL perchloric acid (CAUTION: corrosive, strong oxidizer, use appropriate precautions), and 0.5 mL water.
- In a set of new 1.5 mL tubes, dilute a sample of each resuspended pellet in hydroxyproline buffer to a volume of 200 µL.
NOTE: Dilutions may range from 1:4 to 1:50 of sample:buffer depending on the expected collagen content of the sample; thus some trial-and-error testing may be needed to determine a dilution factor that is appropriate for the given sample set (i.e., place the samples toward the middle of the standard curve). - Prepare hydroxyproline standards. Dilute hydroxyproline in hydroxyproline buffer (see section 7.5.1) to 80 µg/mL. Perform serial dilutions to make 6-8 200 µL standards between 0-20 µg/mL.
- Add 150 µL 14.1 mg/mL Chloramine T solution to each standard and diluted sample. Vortex and incubate at room temperature for 20 min.
- Add 150 µL aldehyde-perchlorate solution to each sample and diluted sample. Vortex and incubate in a heating block at 60 °C for 15 min. Dispose of the excess aldehyde-perchlorate solution as hazardous waste according to local regulations (contains perchloric acid).
- Allow standards and samples to cool, before aliquoting 200 µL of each, in duplicate, into 96-well plates.
- Read plate at 550 nm in a spectrophotometer. Dispose of plate and the remaining volume in 1.5 mL tubes as hazardous waste according to local regulations (contains perchloric acid).
- Calculation of total collagen and collagen fraction.
- Convert the absorbance value for each sample to micrograms of hydroxyproline using the hydroxyproline standard curve.
- Multiply each well by 2.5 to calculate amount of hydroxyproline in the diluted sample. Recall that only 200 of the 500 µL total mixture (200 µL diluted sample + 150 µL chloramine T +150 µL AP solution) is added to each sample well.
- Multiply by dilution factor (Section 7.7) to calculate amount of hydroxyproline in original sample.
- Divide by 0.137 to calculate the amount of collagen (assumes that collagen contains 13.7% hydroxyproline20).
NOTE: Mammalian hydroxyproline abundance in collagen varies slightly across tissues and mammalian species; for example, pig and sheep Achilles tendon contain 13.5 and 13.7% (hydroxyproline mass/dry tissue mass), respectively21. Here, use 13.7% to estimate percent hydroxyproline in collagen, which is used to calculate the collagen content of a tissue sample using the following equation:
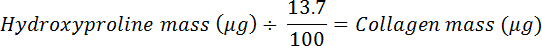
- Divide by the dry mass to calculate the collagen fraction and convert to a percentage.
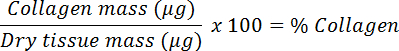
8. Quantification of Molecular Endpoints
NOTE: In addition to primary outcomes of tensile testing and collagen content, molecular endpoints can be measured on 2D or 3D tissue to add mechanistic insight. Bioassays can be used to determine molecular endpoints (see the following section below for context The impact of the post-exercise serum milieu on in vitro ligament function).
- For 3D tissue:
- Prepare constructs as per Step 5 above.
- After construct treatment/intervention, snap-freeze constructs in liquid nitrogen.
- Using a mortar and pestle cooled on dry ice, grind constructs into a powder. Continue at step 8.4/5).
- For 2D tissue:
- Culture human ACL fibroblasts to confluence in a monolayer in six-well plates containing DMEM.
- Aspirate DMEM and apply treatment media according to your experimentation strategy (e.g., time course or dose response experiments).
- Aspirate treatment media and wash cells with PBS.
- Scrape cells to collect using an appropriate extraction buffer/reagent (see next).
- Protein expression analysis: Use a cytosolic extraction buffer (e.g., 250 mM sucrose, 50 mM Tris pH 7.4, 5 mM MgCl2, and protease/phosphatase inhibitor cocktail) to obtain protein lysates. Perform protein concentration assay and continue analysis according to standard western blot procedures.
- Gene expression analysis: Isolate total RNA using 500 µL RNA isolation reagent according to the manufacturer's instructions to obtain high-quality RNA. Perform reverse-transcription and real-time quantitative PCR analysis according to standard procedures.
- DNA isolation: Isolate and quantify genomic DNA using DNA isolation reagent according to the manufacturer's instructions. Quantify DNA concentration by using a spectrophotometer to measure the sample absorbance at 260 nm.
Overview of engineered ligament formation and experimental intervention
Figure 1 shows an overview of the formation of engineered ligaments. Brushite cement, a bone substitute material22 is prepared by combining an orthophosphoric acid/citric acid solution with β-tricalcium phosphate in cylindrical wells. Alternatively, if not directly measuring mechanical function of the tissues, 3-0 silk sutures can be used as anchors in the formation of a tissue. These are pinned 12 mm apart in silicone-coated 35 mm dishes and sterilized by soaking in 70% ethanol. Fibroblasts are isolated from anterior cruciate ligament remnants obtained during ACL reconstruction surgery. After expansion, 2.5 x 105 cells are encapsulated in a fibrin gel formed in the brushite cement anchor-pinned dish. After formation, ligament constructs can be examined for changes in mechanical properties, collagen content, cell proliferation, gene expression, protein levels, and tissue morphology.
Throughout culture, the cells contract the fibrin gel and form a linear tissue between the two anchors (Figure 2A). After 1-2 days in culture, the cells have attached to the fibrin gel, extended cell processes, and started to exert traction forces (Figure 2B). As the fibrin gel is contracted by traction forces and broken down by cellular enzymes, tension is generated between the two anchor points and our cells align parallel to this axis (Figure 2B) and begin to deposit collagen. After 4-5 days, the constructs have contracted around the anchors forming a linear cylindrical tissue (Figure 2A; at this point external stimuli may be applied to the system (intervention at this time avoids disrupting the linear tissue formation process). Interventions may consist of supplementing the culture media with human or animal serum after a given intervention, exogenous cytokines and growth factors, employing mechanical stimulation, or changing other environmental factors such as oxygen tension. Using growth media (DMEM with 10% FBS and 100 U/mL penicillin) supplemented with 200 µM ascorbic acid, 50 µM L-proline, and 5 ng/mL TGF-β1, we have determined that cell proliferation continues throughout a 2 week culture period (Figure 2C) and indeed, light microscopy reveals a dense tissue containing highly aligned cells at 14 days of culture (Figure 2B).
Assessment of engineered ligaments
At the end of the culture period, engineered ligaments can be assessed in a variety of ways. A major advantage of this system is the ability to determine functional changes to the tissue via mechanical testing, a vital assessment given the mechanical role of native ligament. Uniaxial tensile testing can be utilized to measure mechanical properties including load to failure, ultimate tensile strength, and Young's modulus. Viscoelastic properties can also be measured with stress relaxation and creep tests. Figure 3A depicts an engineered ligament held in reverse molded grips in a custom-built uniaxial tensile tester. The right grip is attached to a force transducer to measure the load across the ligament as the tissue is strained to failure. Figure 3B shows a representative stress-strain plot for a test to failure. After undergoing mechanical testing, the same constructs can be dried and processed for a hydroxyproline assay23 to assess total collagen content as well as other biochemical assays. With a sufficient number of additional samples per condition, a thorough examination of an experimental intervention can be conducted, including its effects on cell proliferation, gene and protein expression, and histological morphology. While 14 days is a typical endpoint for our studies, engineered ligaments continue to improve in their mechanical properties and collagen content through 28 days of culture as shown in Figure 3C and can survive for at least 3 months in culture24.
Donor variability is an important consideration for experimental repeatability. Figure 4 shows a representative experiment reported by Lee et al.25 comparing 7 different ACL donors (n = 3 male and n = 4 female) demonstrating typical tensile properties and collagen content after a 2-week culture in the previously described supplemented growth media. Using cells from similar ACL collections, age of donor, time after injury, gender, etc., the engineered ligaments demonstrate low variability between donors and similar characteristics between male and female donors with the exception of the difference in collagen fraction. In the aforementioned study, engineered ligaments were used as an in vitro model to investigate why women have a significantly greater risk of ACL injury than men, and demonstrated that ACL fibroblasts isolated from female donors do not inherently form weaker and less collagenous engineered ligaments25.
The impact of the post-exercise serum milieu on in vitro ligament function
We have previously demonstrated the ability of engineered ligaments to be used to probe physiological processes1,25. In the following representative experiment as reported by West et al.1, we determined the biochemical effects of exercise on ligament function and highlight the methodology and findings here. We formed engineered ligaments using human ACL cells and applied an intervention, at day 7 of culture, consisting of culture media conditioned with human serum collected pre- or post-exercise. Briefly, we recruited healthy young male participants and collected blood samples before and after an acute bout of resistance exercise that increases circulating hormones and cytokines including human growth hormone (GH). Human serum was isolated from pre- and post-exercise blood samples and used in place of fetal bovine serum in the culture media for the second week of engineered ligament culture (Figure 5A). Pre- and post-exercise serum samples were analyzed using ELISA for changes in GH and insulin-like growth factor 1 (IGF-1), the concentration of which can be altered by exercise (Figure 5B). This information was used to correlate serum effects on the engineered ligaments with changes in the serum in response to exercise. After a 14 day culture period, the ligament constructs were evaluated using mechanical testing and a hydroxyproline determination of collagen content and demonstrated a significant increase in both maximal tensile load and collagen content in response to the post-exercise serum. In aiming to assess whether this effect was related to exercise-induced releases of GH or IGF-1, engineered ligaments were formed in a separate experiment and treated with a dose response of either human recombinant GH or IGF-1. Interestingly, while serum GH increased in the blood (Figure 5B-i), progressively increasing recombinant GH concentration in the culture media did not increase collagen content (Figure 5D-i) or mechanical properties (data not shown) in engineered ligaments. In contrast, serum IGF-1 levels did not increase after exercise, but a dose-response experiment revealed that increasing levels in the culture media improved the collagen content of ligament constructs (Figure 4D-ii). Thus, whereas exercise did result in robust increases in post-exercise GH, the dose-response experiment using rhGH raises doubt as to whether GH is directly responsible for the phenotypic enhancement of the engineered ligaments (at least, the 22 kDa isoform alone does not appear to be responsible). Conversely, whereas serum IGF-1 was not altered at 15 min post-exercise, testing rhIGF-1 over a broad range of concentrations revealed that IGF-1 is capable of improving collagen content; however, it should be noted that increasing rhIGF-1 concentrations through a range that estimated physiological levels did not significantly increase collagen content. Thus, the unique post-exercise serum environment was important toward improving the mechanics and collagen of engineered ligaments.
In the study highlighted here1, the volume of experimental serum was limited due to ethical considerations; so, short-term 2D bioassays, which had lower serum demands, were used to further probe the molecular mechanisms responsible for the increase in collagen that was observed. ACL fibroblasts were cultured to confluence in 6-well plates and treated for 1 hour with rest or post-exercise serum, and compared with dose responses of recombinant GH, IGF-1, TGF-β1 and the activation of targets in the PI3K/mTORC1, ERK1/2, and Smad signaling pathways were determined. In the presence of post-exercise serum, the PI3K/mTORC1 and ERK1/2 pathway showed greater activation as assessed by phosphorylation of S6K (Figure 5D-i) and ERK1/2 (Figure 5D-ii), respectively. Compared to the hormone and cytokine dose responses, while GH had a small positive effect on mTOR signaling (Figure 5D-i) and IGF-1 showed a positive effect at the lowest dose, the three treatments of GH, IGF-1, and TGF-β1 did not account for the increase in PI3K/mTORC1 and ERK1/2 signaling. Taken together, our 3D engineered ligament model and 2D bioassay data suggests that the post-exercise serum environment is able to improve engineered ligament function and collagen content through activation of the PI3K/mTORC1 and ERK1/2 pathways.
In summary, using an engineered ligament model combined with exercise-conditioned serum, we were able to i) investigate the effect of post-exercise serum environment on engineered ligament function and collagen, ii) correlate changes in ligament phenotype with changes serum hormone concentration, with the aim to determine which changes in the serum led to changes in engineered ligaments, and iii) further the scope of the work by using 2D bioassays to probe molecular targets of the serum biochemical milieu to determine molecular mechanisms that are activated by post-exercise serum that lead to improvements in ligament function.
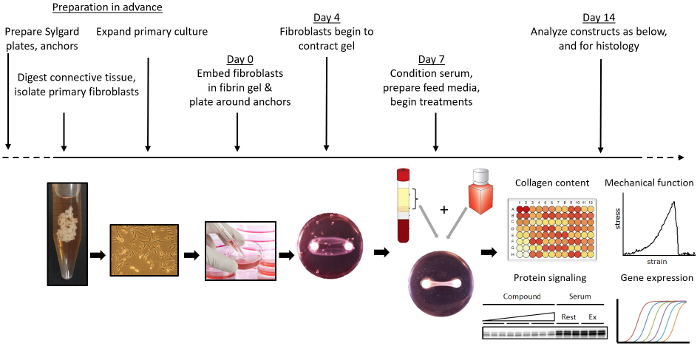
Figure 1: Overview of the formation and use of engineered ligaments. Brushite cement anchors are manufactured and pinned into silicone-coated plates. Primary fibroblasts are isolated and expanded from ACL remnants. Engineered ligaments are formed by encapsulating fibroblasts in a fibrin gel around two brushite cement anchors. Engineered ligaments are cultured and treated with whichever specific chemical or mechanical (e.g., via a bioreactor) stimuli desired. At the desired endpoint, engineered ligaments can be collected and assessed for mechanical properties, gene expression, collagen content, protein expression, and histology. Please click here to view a larger version of this figure.
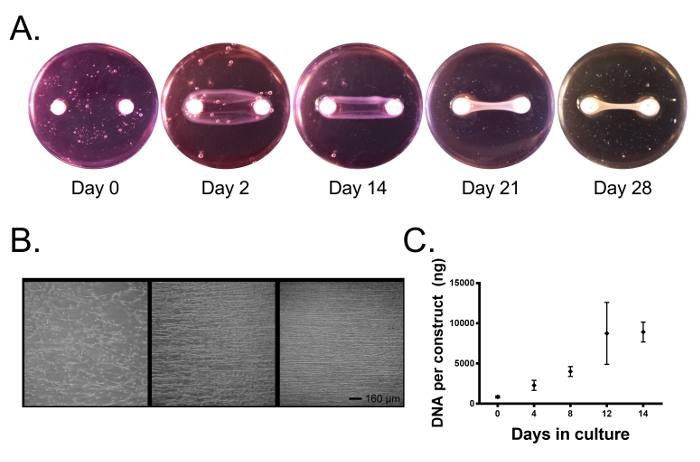
Figure 2. Primary ligament fibroblasts form a fibrin-based bone-to-bone engineered ligament spanning two brushite cement anchors. (A) Over time, the fibroblasts contract the fibrin gel around the brushite cement anchors forming a linear tissue. (B) In the first three days, the cells attach to the fibrin gel and exert traction forces, aligning the cells with the long axis of the construct. Over 14 days, the cells form a highly aligned tissue. Scale bar = 160 µm. (C) The DNA content of the engineered ligaments continues to increase over 14 days in culture as the cells proliferate. Data is presented as mean ± SD with n = 3-4 constructs per group. groups. Please click here to view a larger version of this figure.
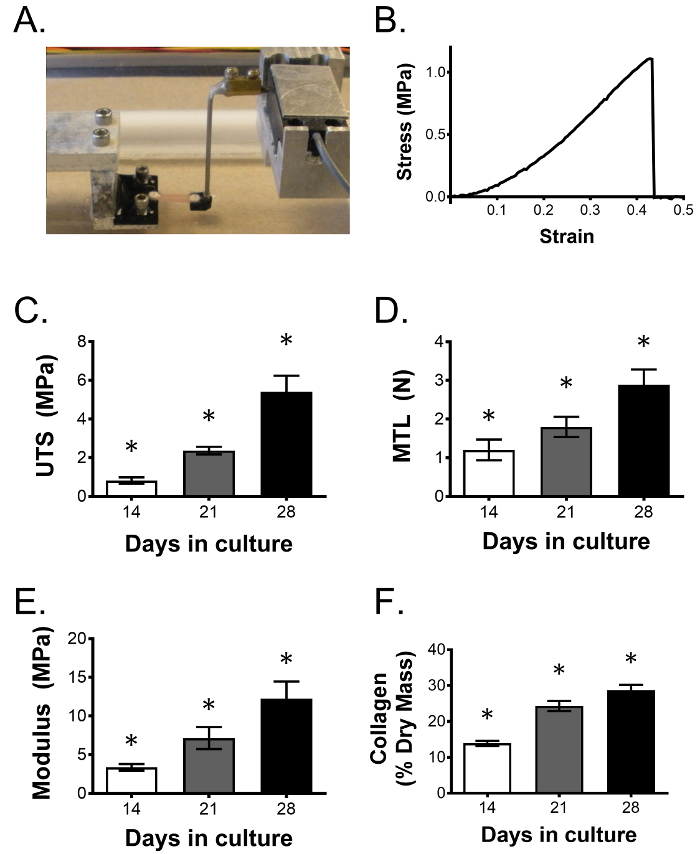
Figure 3: Engineered ligament mechanical properties and collagen content improve over time. (A) Ligament constructs are uniaxially tensile tested to determine the effect of a given intervention on ligament function. As shown, two reverse molded, 3D printed grips hold reciprocally shaped anchors that are bridged by the engineered sinew. Anchors are coupled to a stepper motor and force transducer to generate stress/strain curves of the tested tissue, allowing mechanical properties can be determined. (B) Representative stress-strain curve from an engineered ligament strained to failure. Over the course of 28 days, (C) ultimate tensile strength (UTS), (D) maximal tensile load (MTL), (E) Young's modulus, and (F) collagen fraction continue to improve. Data are presented as mean ±SD with n = 5 constructs per group. * indicates significant difference from all other groups. Please click here to view a larger version of this figure.
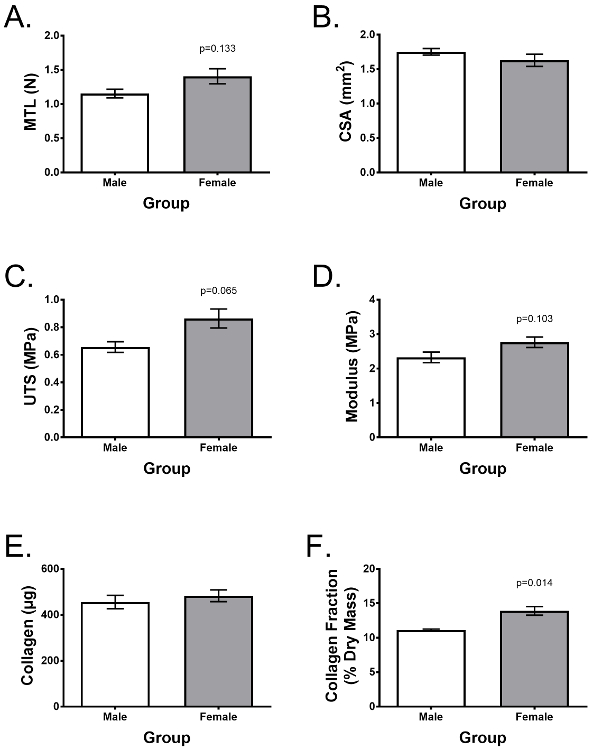
Figure 4: Engineered ligaments can be assessed for functionality and biochemical content, displaying low donor variability. Engineered ligaments were formed 7 different donors (n = 3 male, n = 4 female). After 2 weeks of culture, they were assessed for differences in (A) maximum tensile load, (B) ultimate tensile strength (UTS), (C) Young's modulus, (D) cross-sectional area (CSA), (E) total collagen content per construct, and (F) collagen as a fraction of dry mass. Data are presented as mean ± SD and statistical significance was with Student's t-test. * indicates significant difference from other groups (p <0.05). Figure adapted from Lee et al.25 Please click here to view a larger version of this figure.
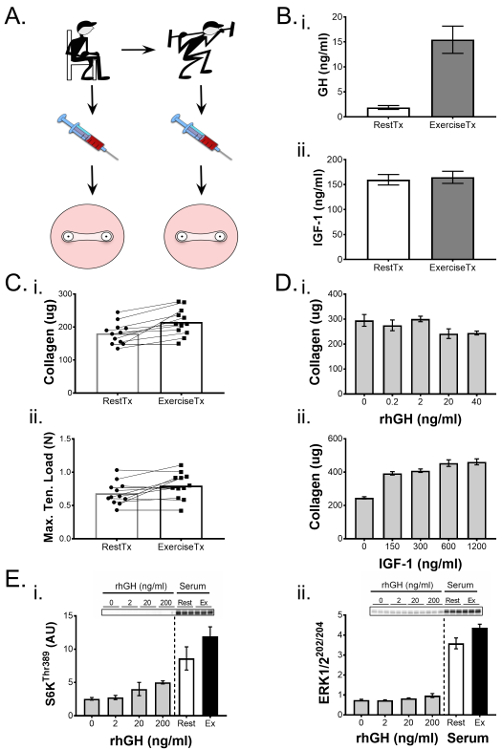
Figure 5. Engineered ligaments demonstrate mechanical and biochemical changes in response to biological interventions. (A) Serum was isolated from blood draws collected from subjects pre- (RestTx) and post-exercise (ExTx) and used to treat engineered ligaments during the second week of culture. (B) (i) Human growth hormone (GH) and (ii) insulin-like growth factor (IGF)-1 levels in the rest and exercise serum were quantified through ELISA. (C) Engineered ligaments treated with ExTx demonstrated improved (i) collagen content and (ii) maximal tensile load. Statistical significance of paired comparisons (RestTx and ExTx) was analyzed by a t-test with significance level set at p <0.05. (D) A dose-response of (i) GH and (ii) IGF-1 was used to determine possible contributions of these factors to the changes in collagen content due to ExTx. E) 2D bioassays were used to compare the effects of increasing doses of GH, RestTx, and ExTx on molecular signaling targets such as the phosphorylation of (i) S6KThr389 and (ii) ERK1/2Thr202/Tyr204. Statistical comparison of more than two experimental groups was performed using ANOVA and Tukey's HSD. Data are presented as presented as mean ± SD. * indicates a significant difference from control (p <0.05) and § indicates significant difference from 150 ng/mL and 300 ng/mL IGF-1. Figure adapted from West et al.1 Please click here to view a larger version of this figure.
