En vanlig egenskap hos störningar och sjukdomar i centrala nervsystemet (CNS), såsom traumatisk hjärnskada (TBI), ryggmärgsskada (SCI), stroke, Alzheimers sjukdom och Parkinsons sjukdom är avbrytandet av axonala vägar och neuronal cell Förlust 1 , 2 , 3 , 4 , 5 , 6 . När exempelvis en ischemisk stroke går obehandlad uppskattas det att axoner förloras med en hastighet av 7 miles axon per minut 5 . När det gäller TBI, som ungefär 1,7 miljoner människor upplever varje år i USA ensam, kan axonal degeneration fortsätta att inträffa år efter trauma, eftersom den initiala skaden utfäller ett långtids neurodegenerativt tillstånd 4 . Försvårande av dessa skadliga effekter har CNS en kraftigt begränsad capaStad för regenerering 1 , 7 , 8 , 9 . Efter skada utvecklas en hämmande miljö som kännetecknas av brist på riktad vägledning till avlägsna mål, närvaron av myelinhämmande hämmare som hindrar neuritutväxten och bildandet av ett glialär av reaktiva astrocyter 8 , 10 , 11 , 12 . Glialärret tjänar som en biokemisk och fysisk barriär mot regenerering, med molekyler som kondroitinsulfatproteoglykaner som hindrar axonutväxten 8 , 11 . Dessutom, även om neurala stamceller har hittats i vuxna CNS, är produktionen av nya neuroner begränsad, eftersom konsekventa bevis på neurogenes endast har hittats i luktlampan, den hippocampalaSubgranulär zon, periventrikulärt område och ryggmärgs 13 , 14 centrala kanal. Dessa hinder hindrar funktionell återhämtning av förlorade neuroner och vit materia arkitektur efter skada eller sjukdom, vilket resulterar i de ofta livsförändrande och långvariga effekterna av dessa tillstånd.
Trots bristen på regenerativ kapacitet hos vuxna CNS har det visat sig att axonal regenerering är möjlig om adekvata miljöanpassningar presenteras för värdvärdena neuroner 15 , 16 , 17 , 18 . Forskare har försökt leverera och manipulera tillväxtfaktorer ( t.ex. nervtillväxtfaktor, epidermal tillväxtfaktor, glialberoende tillväxtfaktor och neurotrofisk faktor-3) och andra vägledningsmolekyler för att stimulera plasticitet och axonregenerering 14 , </ Sup> 18 , 19 . Trots att dessa studier har bekräftat att vuxna axoner är kapabla att reagera på tillväxtfaktorer begränsas dessa strategier av lågt permeabilitet hos blod-hjärnbarriären och de specifika rumsliga och temporala gradienterna som krävs för att främja regenerering 14 , 18 , 19 . Andra tillvägagångssätt har åberopat hyperaktivering av regenerationsrelaterade transkriptionsfaktorer i CNS-neuroner. Till exempel stimulerade överuttryck av stat3-transkriptionsfaktorn axonal regenerering i optisk nerv 20 . Ändå misslyckas både biomolekylavgivning och överuttryck av transkriptionsfaktorer att ersätta förlorade neuronpopulationer. Cellbaserade strategier har huvudsakligen koncentrerat sig på transplantation av neurala stamceller (NSC) i CNS, med fördel av deras förmåga att ersätta CNS-neuroner, frigöra trofiska faktorer,Och stödja försöken på neurogenes som uppträder efter skada 17 . Trots detta finns det fortfarande pressande utmaningar som hindrar detta tillvägagångssätt, inklusive de transplanterade neurala cellernas hämmade förmåga att överleva, integrera med värden och förbli spatialt begränsad till det skadade området 6 , 14 , 17 , 21 . Dessutom är celltillförsel enbart oförmögen att återinföra cytoarkitekturen av skadade eller förlorade axonala vägar. Ett alternativt tillvägagångssätt som behandlar problemen med cell- och läkemedels- och kemikalieleveransstrategier är att kombinera dessa metoder med användningen av biomaterial 14 , 22 , 23 . Biomaterial som hydrogeler kan emulera de biokemiska och fysikaliska egenskaperna hos den extracellulära matrisen (ECM), som hjälper till vid celltillförsel enD retention inom det skadade området och leverera tillväxtfaktorer och andra bioaktiva molekyler med kontrollerat frisättning 22 . De attraktiva egenskaperna hos dessa biomaterialbaserade strategier har resulterat i bevis på in vivo axonal regenerering efter transplantation av byggnadsställningar till det belastade området 24 , 25 , 26 , 27 , 28 , 29 , 30 . Emellertid ersätter acellulära biomaterialstrategier inte förlorade neuronala populationer; När de används som leveransfordon för neuron-, glial- eller neuronprekursorceller, är biomaterialen inte kapabla att rekonstituera axonala nätverk på lång avstånd. Utmaningen att utveckla ett tillvägagångssätt som tacklar både den axonala vävnadsdegenerationen och neuronförlusten i samband med CNS-skada och sjukdom kvarstår <Sup class = "xref"> 31.
Vår forskningsgrupp rapporterade tidigare utvecklingen av implanterbara mikrovågsmetallerade neurala nätverk (mikro-TENN), som är en typ av "levnadsställare" som består av neuroncellceller som är bundna till en eller båda ändarna av en agaroshydrogel-ECM-mikrokolonn , Med orienterade axonala kanaler som sträcker sig genom det inre av denna tredimensionella (3D) inneslutning 1 , 10 , 31 , 32 . En av de största skillnaderna mellan denna teknik och tidigare tillvägagångssätt är att cytoarkitekturen av mikro-TENN är skapad helt in vitro och transplanteras efteråt 33 , 34 , 35 , 36 , 37 , 38 , <Sup class = "xref"> 39 , 40 , 41 . In vitro- tillverkning erbjuder omfattande rumslig och tidsmässig kontroll av cellulär fenotyp och orientering, mekaniska / fysikaliska egenskaper, biokemiska signaler och exogena faktorer, vilket gynnar integrationen av dessa byggnadsställningar med värden efter implantationen 41 , 42 . Mikro-Tenn är anatomiskt inspirerade eftersom de emulerar neuroanatomi i hjärnan, som visar axonala områden som liknar de som överbryggar distinkta funktionella regioner i hjärnan ( Figur 1A ) 1 . Därför kan denna strategi kunna fysiskt ersätta förlorade vita substansområden och neuroner efter implantation till en belastad region. Denna teknik är också inspirerad av utvecklingsmekanismer där "naturliga levnadsställningar" som bildas av radiella glialceller och banbrytande axoner fungerar som vägleder guider för cellMigration från subventrikulär zon och axonal utväxt, respektive 43 . Dessa mekanismer återfinns i de anpassade axonala områdena av mikro-TENN, som kan presentera levande vägar för neural cellmigration och axonal regenerering genom axonmedierad axonal utväxt ( Figur 1C ) 43 . Vidare utnyttjar denna strategi synaptisk integration mellan mikro-TENN-neuronerna och de inhemska kretsarna, som bildar nya reläer som kan bidra till funktionell återhämtning ( Figur 1B ) 43 . Kapaciteten för synapsbildning kan också ge detta tillvägagångssätt förmågan att modulera CNS och reagera på värdvävnad enligt nätverksåterkoppling. Till exempel kan optogenetiskt aktiva neuroner i de levande ställningarna stimuleras för att modulera värdneuroner genom synaptiska interaktioner ( Figur 1D ).
Dessutom är den biomaterialbaserade rörformigaUgn av mikro-TENN erbjuder en lämplig miljö för celladhesion, tillväxt, neuritillägg och signalering, medan konstruktionens miniatyrdimensioner möjliggör minimalt invasiv implantering och ger en delvis sekvestrerad mikromiljö för gradvis integration i hjärnan. Faktum är att senaste publikationer har visat potentialen hos mikro-tenn att efterlikna neurala vägar efter implantation i råtthjärnan. Efter stereotaxisk mikroinjektion rapporterade vi tidigare bevis på mikro-TENN-neuronal överlevnad, underhåll av axonaltraktionsarkitektur och neuritillförsel i värdcortexen till minst 1 månad in vivo 10 , 31 . Vidare gav märkning med synapsin histologiska bevis på synaptisk integration med nativ vävnad 10 , 31 . Sammantaget kan mikro-tenn vara unikt anpassade för att rekonstruera och modulera skadadeCNS genom att ersätta förlorade neuroner, synaptiskt integrera med värdkretsen, återställa förlorad axonal cytoarkitektur och, i vissa fall, tillhandahålla regenererande axoner med lämpliga sökvägsljus.
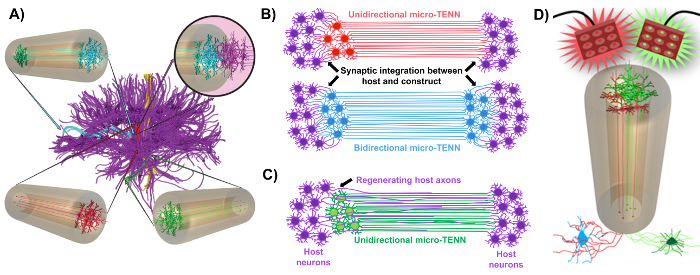
Figur 1: Principer och inspiration bakom utvecklingen av mikrovävnadstekniska neurala nätverk (mikro-TENN). ( A ) Micro-TENNs efterliknar cytoarkitekturen i hjärnans anslutning (lila), i vilken funktionellt distinkta regioner är kopplade med långa, inriktade axonala områden i enriktad (röd, grön) eller dubbelriktad (blå) väg. Till exempel kan mikro-tenn rekonstruera förlorade förbindelser i kortikotalamiska och nigrostriatala vägar eller i perforantvägen från entorhinal cortex till hippocampus (anpassad från Struzyna et al. , 2015) 1 . ( B ) Diagram över en enhetlig riktningL och dubbelriktad mikro-TENN (röd respektive blå) synaptiskt integrera med värdkretsen (lila) för att fungera som ett funktionellt relä mellan båda ändarna av en lesion. ( C ) Schematisk av axonaltraktionerna i en enriktad mikro-TENN (grön) som tjänar som en vägledning för axon-underlättad regenerering av värdaxoner (lila) mot ett mål som mikro-TENN interagerar med. ( D ) Konceptuellt diagram över användningen av optogenetiskt aktiva mikro-TENNS som neuromodulatorer, utnyttja synaptisk integration med excitatoriska eller hämmande neuroner (botten). Vänligen klicka här för att se en större version av denna figur.
Det nuvarande manuskriptet beskriver den metod som används för att tillverka mikro-TENN med användning av embryonellt härledda cerebrala kortikala nervceller. Speciellt kan mikro-tenn tillverkas med andra typer av neurala celler. För exRiklig, de första rapporterna om framgångsrik mikro-TENN-utveckling presenterade dorsala root ganglion (DRG) neuroner 32 . Hydrogelmikrokolumnerna kan alstras ( Figur 2A ) genom att tillsätta flytande agaros till en skräddarsydd, laserskärd cylindrisk kanalgrupp eller till kapillärrör, vilka båda innehåller inriktade akupunkturnålor. Nålen bildar lumen och bestämmer mikrokolonnens innerdiameter (ID), medan kapillärrörets ID och diametern hos cylindrarna i den laserskärda anordningen dikterar konstrukternas ytterdiameter (OD). OD och ID kan väljas enligt önskad applikation genom att välja olika diametrar för anordning / kapillärrör respektive akupunkturnålar. Längden på mikrokolumnerna kan också varieras; Hittills har vi rapporterat konstruktionen av mikro-tenn upp till 20 mm i längd 10 och arbetar aktivt med längre längder. Efter agarosgelerna och akupunkturen nEedlar avlägsnas, en ECM-lösning som i allmänhet består av kollagen av typ I och laminin tillsätts till konstruktionens lumen ( figur 2C ). ECM-kärnan ger en byggnadsställning som stödjer neuronal celladhesion och axonal utväxt. Ursprungligen pläterades primära råtta-kortikala neuroner i mikrokolonnerna med användning av dissocierade cellsuspensioner 10 , 31 , 32 . Detta tillvägagångssätt producerade emellertid inte målcytoarkitekturen i alla fall, som definierades som neuroncellcellerna begränsade till ändarna av mikrokolumnerna, varvid den centrala lumen bestod av rena inriktade axonala kanaler. Sedan dess har användningen av en tvungen neuronal aggregeringsmetod (baserat på protokoll anpassade från Ungrin et al .) Möjliggjort en mer pålitlig och konsekvent tillverkning av mikro-TENN med den ideala strukturen ( Figur 2B ) 44 . Förutom att beskriva strömmenMetodik, kommer denna artikel att visa representativa faskontrast och konfokala bilder av mikro-TENN som visar bildandet av axonala kanaler över tiden, såväl som den slutliga målcytokarchitecturen. Detta manuskript kommer också att expandera på anmärkningsvärda aspekter av protokollet och återstående utmaningar och framtida riktningar för mikro-TENN-tekniken.
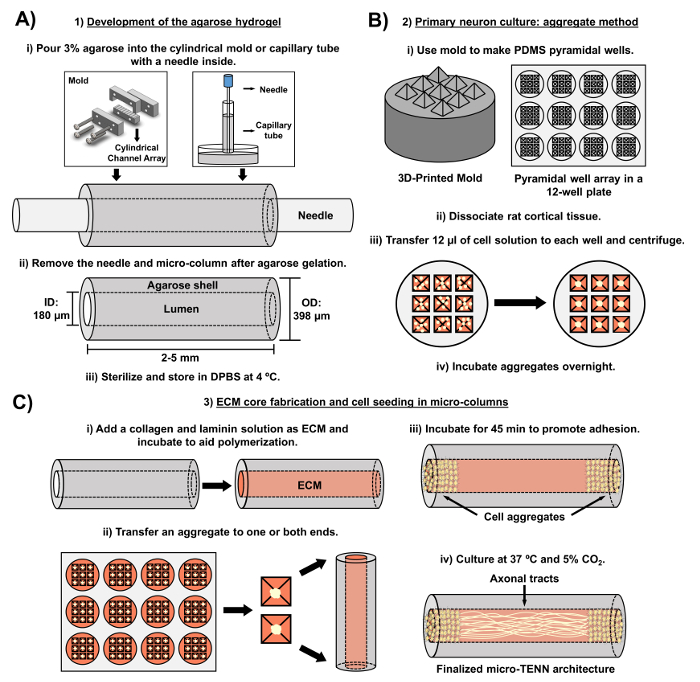
Figur 2: Schematiskt diagram över tre-stegs mikro-TENN-tillverkningsprocessen. ( A ) Utveckling av agaroshydrogelen: (i) Initialt införs en liten akupunkturnål ( t.ex. 180-350 um diameter) i de cylindriska kanalerna av en skräddarsydd, laserskärd form eller ett kapillärrör ( t.ex. , 380-700 pm i diameter). I nästa steg införs flytande agaros i DPBS i de cylindriska kanalerna eller kapillärrören. (Ii) Efter agarosgelerna avlägsnas nålen ochFormen avmonteras för att ge de ihåliga agarosmikolonnerna. (Iii) Dessa konstruktioner steriliseras sedan och lagras i DPBS. ( B ) Primär neuronkultur och aggregatmetoden: (i) Neuronal aggregering utförs i pyramidala mikrobrunnsarrayer, gjutna av 3D-tryckta formar, som passar in i brunnarna i en 12-brunns odlingsplatta. (Ii) Mikro-TENN inkluderar primära råttneuroner dissocierade från fostrets hjärnor av embryonala-18-råttor. Efter vävsdissociation med trypsin-EDTA och DNas I framställs en celllösning med en densitet av 1,0-2,0 x 106 celler / ml. (Iii) 12 | il av denna lösning överförs till varje brunn i den pyramidala mikrobrunnmatrisen. Plattan innehållande dessa mikrobrunnar centrifugeras för framställning av cellaggregat. (Iv) Dessa inkuberas sedan över natten före plätering i mikrokolonnerna. ( C ) ECM-kärntillverkning och cellsåning: (i) Före cellsåkning fick en ECM-lösning innehållande 1 mg / ml typ I-kollagen och 1 mg / mlLaminin överförs till det inre av mikro-tenn och tillåts polymerisera. (Ii) Beroende på huruvida enriktad eller dubbelriktad mikro-TENN tillverkas, placeras ett aggregat vid respektive ena eller båda extremiteterna av mikrokolonnen. (Iii) Efter en inkubationsperiod för att främja vidhäftning odlas mikro-TENN i petriskålar som översvämmas med kompletterat embryonalt neuronalt basalt medium. Iv) Efter 3-5 dagar i odling bör den slutliga mikro-TENN-strukturen demonstrera cellaggregat vid ytterkolumnerna, med axonala kanaler som sträcker sig över längden. Vänligen klicka här för att se en större version av denna figur.
