Human lung tumors are usually highly infiltrated in CD8 T cells that are preferentially found in the stroma3,4. By following this protocol one should expect to visualize a large number of immunostained CD8 T in human lung tumor slices. A co-labeling with anti-EpCAM and anti-CD90 antibodies should permit to delineate tumor islets and stromal areas. Fibrillar collagen, abundant in the tumor stroma, can be visualized without exogenous staining by using second harmonic generation on a two-photon microscope3,4. Note that some human lung tumors present no clear distinction between tumor islets and stroma. Such tumors should be discarded as well as specimen presenting large areas of cell necrosis identified by non-specific binding of antibodies and DAPI staining. Consistent with published results obtained in human lung and ovarian carcinomas, resident CD8 T cells are more concentrated in the stroma compared to tumor islets4. A representative example of CD8 T cell distribution in relation to tumor cells and CD90 is shown in Figure 2.
The slice assay allows for the analysis of resident T cell motility in different tumor regions. Useful criteria for comparing CD8 T cell motility in different regions include the average velocity (path length divided by the time duration sampled) and the displacement length (the vector between the beginning and end of a track). Although scarce in tumor islets, CD8 T cells migrate more actively in this compartment compared to the stroma (Figure 3). On average, the mean velocity of individual CD8 T cells should be close to 4 µm/min in the tumor stroma with high intra and inter-tumor variabilities4. In the tumor stroma, the motility of resident T cells is strongly influenced by the density and orientation of extracellular matrix fibers3,4. A successful experiment will result in less T cells present in dense matrix areas compared to loose matrix regions, consistent with published results3,4.
Note that some human lung tumors exhibit a very high level of autofluorescence precluding imaging of resident T cells. Such feature is rapidly observed at the beginning of the imaging session. Autofluorescent tumors should be discarded. A representative example of resident CD8 T cells in an autofluorescent tumor is shown in Figure 4.

Figure 1: Photographs of the perfusion system. A) The phenol red-free RPMI solution is enriched in 5% C02, 95% O2. B) The solution is then perfused by a peristaltic pump. C) The RPMI solution enters the culture dish via the inlet. At the outlet, the solution is aspirated by the pump from the chamber into a waste container. Note that the aspiration tip is set at a level to determine solution height. Please click here to view a larger version of this figure.
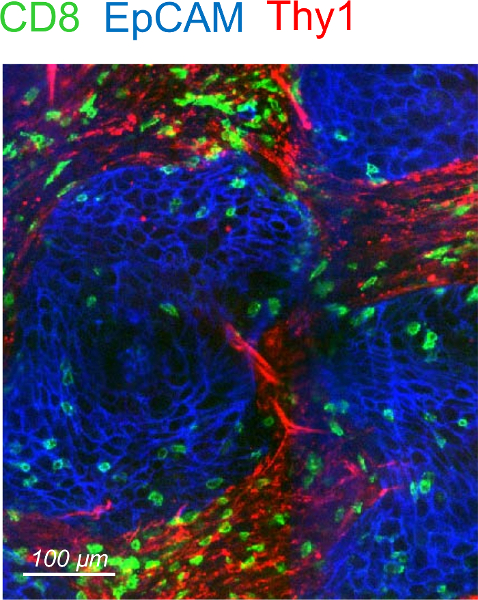
Figure 2: Distribution of resident CD8 T cells in a human lung tumor. Representative image of a human lung tumor slice stained for CD8 (green), EpCAM (blue) and CD90 (red). Please click here to view a larger version of this figure.
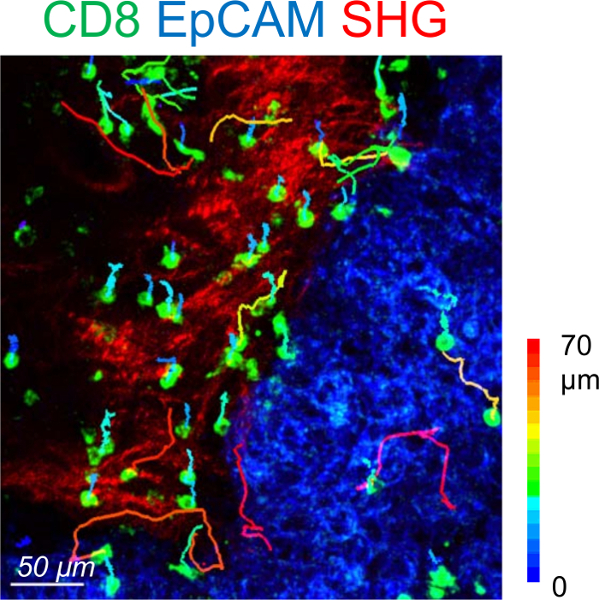
Figure 3: Motility of resident CD8 T cells in a human lung tumor. Trajectories of individual resident CD8 T cells in the stromal (red) and tumor cell (blue) regions of a human lung carcinoma slice. The slice was stained with the indicated antibodies and imaged for 20 min with a confocal microscope. The fibrillary collagen was detected at the end by using second harmonic generation (SHG) on a two-photon microscope. Tracks are color-coded according to CD8 T cell displacement length. This image has been modified from 4. Please click here to view a larger version of this figure.
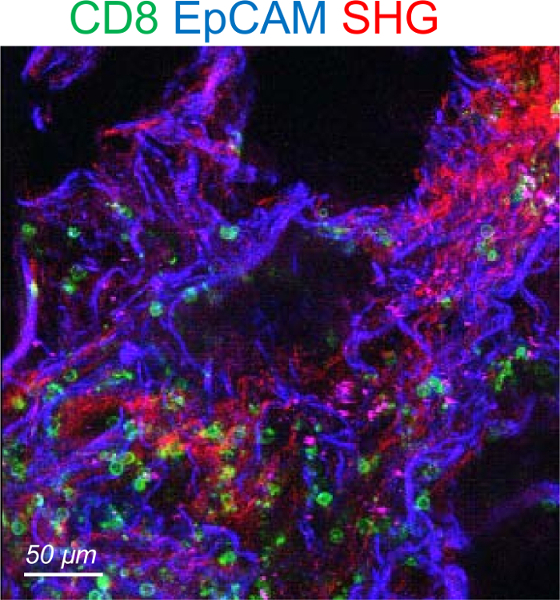
Figure 4: Representative image of an autofluorescent human lung tumor. The tumor slice was stained for CD8 (green) and EpCAM (blue). The SHG signal is in red. Please click here to view a larger version of this figure.
