وتؤدي الميكروبات المرتبطة بالمصانع أدوارا هامة في ركوب الدراجات الكيميائية الجيولوجية الحيوية، والمعالجة البيولوجية، والتخفيف من تغير المناخ، ونمو النباتات والصحة، وتسامح النبات للإجهادات الأحيائية وغير الحيوية. الكائنات الحية الدقيقة تتفاعل مع النباتات مباشرة من خلال الخلايا النباتية الجدار الاتصال وبشكل غير مباشر عن طريق إفراز الكيميائية والإشارات 1 ، 2 ، 3 . وبوصفها كائنات حية، وضعت النباتات آليات مباشرة وغير مباشرة لمقاومة العدوى بواسطة مسببات الأمراض. وتشمل الدفاعات المباشرة الدفاعات الهيكلية والتعبير عن بروتينات الدفاع، في حين أن الدفاعات غير المباشرة تشمل إنتاج المستقلب النبات الثانوي وجذب الكائنات الحية معادية للغزو مسببات الأمراض 4 ، 5 . الجذور المستمدة من الافرازات الجذر، إفرازات، ميوسيلاجيس، موسيجيل، و ليسيتس يغير الخصائص الفيزيائية والكيميائية لل ريزوسفير لجذب أو صدالميكروبات نحو المضيفين 6 . التركيب الكيميائي لإفراز الجذر هو نوع معين، وبالتالي بمثابة مرشح انتقائي يسمح بعض الكائنات الحية الدقيقة قادرة على الاعتراف هذه المركبات لتزدهر في الجريزوسفير 6 . وهكذا، يمكن تحفيز الأنواع الميكروبية المتوافقة لتنشيط وتعزيز جمعياتها، إما لصالح أو ضرر للمضيف النبات 1 .
ويعد فهم تفاعلات الميكروب النباتي في الغلاف الجوي أمرا أساسيا لتعزيز إنتاجية النبات وأداء النظام الإيكولوجي، حيث أن غالبية التعرض للميكروبات والكيماويات يحدث في البنية الجذرية والسطح البيني للتربة والهواء 2 و 6 و 7 و 8 . ومع ذلك، فإن فحص التفاعلات بين النباتات الميكروبية تحت الأرض والاستجابات المتبادلة كان تحديا بسبب ما يثير الفضول والطبيعة المعقدة والديناميكية وعدم وجود نماذج تجريبية مناسبة مع هيكل الجذر الطبيعي ومورفولوجيا النبات تحت ظروف نمو يمكن السيطرة عليها بإحكام. وباعتبارها واحدة من أكثر النباتات النباتية التي تمت دراستها بشكل كبير، فإن الجراثيم تصيب مجموعة واسعة من النباتات ذات الأهمية الزراعية والبستانية، بما في ذلك الكرز والتفاح والكمثرى والعنب والورود 9 . الجراثيم هو كائن نموذج مهم لفهم التفاعلات الممرضات النباتية وهو أداة قوية في تحويل النبات والهندسة النباتية 10 ، 11 ، 12 ، 13 ، 14 .
وقد تم دراسة التفاعلات الجراثيم دراسة جذور لعدة عقود، والفهم الحالي للأمراض الجراثيم المرضية واسعة النطاق 9 ،f "> 11 ، 15 ، 16. ويعزى المرضية الجرثومية إلى حد كبير إلى قدراتها المتطورة لإدراك الإشارات المستمدة من النباتات، مما أدى إلى التشكيل الدقيق لبرنامج الفوعة والاتصالات من خلية إلى خلية، ما يسمى استشعار النصاب 17 . وينظم برنامج الجرثومة الفوعة من قبل العديد من الإشارات المتاحة في ريزوسفير وينطوي على مجموعتين من أنظمة 2-مكون، ونظام تشفغ / I ونظام فيرا / G.الظروف الحمضية في ريزوسفير تفعيل النسخ من تشفغ / I ، فيرا / G ، والعديد من الجينات الأخرى المشاركة في الأجرعية المرضية، بما في ذلك virE0، virE1، virH1، virH2، والجينات من نوع VI نظام إفراز (T6SS) 18. المركبات الفينولية، بما في ذلك acetosyringone (4'-هيدروكسي-3، 5 مصنع مشتقة '-dimethoxyacetophenone)، وتفعيل Vإيرا / G 2-نظام مكون من خلال آليات الإشارات الفسفرة 19 . فيرا / G ثم ينشط ريجيونون الفيروس بأكمله، مما أدى إلى نقل والتكامل من ~ 20 كيلو بايت جزء الحمض النووي البكتيرية تسمى الحمض النووي نقل (T- دنا) من البلازميد (تي) التي تحفز الورم في نواة النبات 16 . T-دنا يحمل الجينات المسؤولة عن تخليق هرمونات النبات إندول -3 حمض الخليك (إيا) ( إام و إاه ) والسيتوكينين ( إيبت )، وعبر مرة واحدة في الخلايا النباتية، ويتم إنتاج كميات كبيرة من هذه الفيتوهورمونات. وهذا يؤدي إلى انتشار الأنسجة غير طبيعي وتطور الورم النباتي، والمعروفة باسم مرض غالون تاج، وهو مشكلة مزمنة وعاد للنباتات 9 ، 11 ، 20 . إيا أيضا يعمل بشكل جماعي مع حمض الصفصاف وحمض الأمينية غاما الأمينية لقمع الفوعة الجرثومية أو للحد من أغروباكتيريو (كور) الاستشعار عن بعد (قس) 17 ، 21 ، 22 . لمكافحة هذا القمع، T- الحمض النووي يحمل أيضا الجينات لالبصرية الحيوي، الذي ينشط أغروباكتريوم النصاب الاستشعار عن تعزيز إمبروكتريوم المرضية وأيضا بمثابة مصدر المغذيات للمسبب 22 ، 23 .
وعلى الرغم من فهم عميق العام للالأجرعية التفاعلات -plant والناتجة نقل T-DNA إلى المضيف النبات، والأحداث الإشارات المعقدة في المرحلة الأولى من التفاعل وفهم أقل أيضا. ويرجع ذلك جزئيا إلى القيود المفروضة على النهج التقليدية للتحقيق الجراثيم- زرع إشارة. وتستخدم الخلايا النباتية تعليق الثقافات والجروح اصطناعية محددة الموقع عادة لدراسة الجزيئية التفاعلات النباتية ميكروب 24 ،إف "> 26 ، 27. ومع ذلك، تعليق خلية تفتقر النموذجية مورفولوجيا النبات، على وجه الخصوص، وخلايا تعليق النبات ليس لديهم هياكل الجذور ونضح الافرازات، والتي هي مهمة جدا لتفعيل الكيميائي الميكروبي والفوعة 28 ، 29. الحفاظ على مورفولوجيا النبات وقد تم معالجة هيكل الجذر عن طريق اصابة النباتات اصطناعيا، مما يسهل العدوى موقع معين، مما أدى إلى الكشف عن الجينات ذات الصلة دفاع النبات التي يسببها في الأنسجة النباتية المصابة مباشرة 30 ، 31. ومع ذلك، الإصابات الاصطناعية يختلف اختلافا كبيرا عن العدوى الممرض في الطبيعة ، لا سيما أن الجرح يؤدي إلى تراكم حمض جاسمونيك (جا)، والذي يتداخل بشكل منهجي مع إشارات النباتات الطبيعية والدفاع 26. وبالإضافة إلى ذلك، وتستخدم المواد الكيميائية الاصطناعية عادة لتحفيز اصطناعية ردود المضيف النباتأو فيروسة الممرض. على الرغم من أن تكملة هذه المركبات الكيميائية تعكس تركيزات في بلانتا ممكن، فإن مثل هذه المكملات لا تمثل نشر الافرازات الجذر تدريجيا في ريزوسفير المحيطة، الذي يولد التدرج الكيميائي الاستشعار عن طريق الميكروبات 28 ، 32 . ونظرا للقيود المفروضة على النهج التقليدية لدراسة تفاعلات الميكروبات النباتية، قد تكون دقة وعمق البيانات التي تم الحصول عليها معوقة ومقيدة، والمعرفة الناتجة عن النهج التقليدية قد لا تترجم مباشرة في النباتات . العديد من جوانب النباتات الزراعية إشارة ليست مفهومة تماما، لا سيما في المرحلة المبكرة من التفاعلات، عندما لم تظهر أعراض المرض بعد.
ولتعديل القيود على النهج التقليدية، يعرض هذا العمل الزراعة المائية غير المكلفة والمتحكم فيها بشكل صارم والمرننظام أوكولتيفاتيون الذي يسمح للباحثين للحصول على رؤى أعمق في مسارات التشوير والاستجابة المعقدة في المرحلة الأولى من التفاعلات الجزيئية النباتية والميكروب. وقد استخدمت الزراعة المائية على نطاق واسع لدراسة المغذيات النباتية، الافرازات الجذر، وظروف النمو، وآثار السمية المعدنية على النباتات 33 ، 34 . وهناك العديد من المزايا للنماذج المائية، بما في ذلك المتطلبات المكانية الصغيرة، وإمكانية الوصول إلى الأنسجة النباتية المختلفة، والرقابة المشددة على العناصر الغذائية / البيئية، ومكافحة الآفات / الأمراض. كما أن أنظمة الزراعة المائية أقل تقييدا لنمو النباتات مقارنة بتقنيات طلاء أجار / فيتواغار التي تقيد النمو عادة بعد 2-3 أسابيع. الأهم من ذلك، والحفاظ على هياكل المصنع كله يسهل إفراز الجذر الطبيعي الضروري للالعلاج الكيميائي الميكروبي والفوعة الحث 8 ، 29 . وصف النظامالسرير هنا هو أبسط وأقل كثيفة العمالة من البدائل 33 ، 34 . ويستخدم أجزاء أقل ولا يتطلب أي أدوات أخرى من مقص القياسية. ويستخدم شبكة معدنية (على عكس النايلون 33 ) كدعم قوي لنمو النبات وطريقة بسيطة من التهوية تحت ظروف معقمة من خلال الهز لدعم النمو الميكروبي. وبالإضافة إلى ذلك، يمكن للنظام استخدام شبكة معدنية من مختلف الأحجام لدعم نمو النبات، الذي يستوعب أنواع النباتات المختلفة دون تقييد عرض جذورها.
في النظام كوكولتفاتيون المائية المعروضة هنا، تزرع النباتات في نظام الزراعة المائية العقيمة حيث جذور النباتات تفرز المركبات العضوية التي تدعم نمو البكتيريا الملقحة. في هذا النظام كوكولتيفاتيون، لا تكمل أي مواد كيميائية اصطناعية، مثل هرمونات النبات، داعية إليتور، أو المواد الكيميائية التي تسبب الفوعة، مما يعكس الخلية الطبيعيةوالتوازن -signaling خلال التفاعلات مصنع الميكروب. مع هذا النظام كوكولتفاتيون المائية، كان من الممكن تحديد في وقت واحد التعبير الجيني في أرابيدوبسيس ثاليانا كول -0 أنسجة الجذر على العدوى من قبل أغروباكتريوم ، فضلا عن تفعيل الجينات الجراثيم على كوكولتيفاتيون مع أرابيدوبسيس . وقد تجلى أيضا أن هذا النظام هو مناسبة لدراسة مرفق الأجرعية إلى جذور النباتات، وكذلك التعريف secretome جذور النبات، على استنبات مشترك (العدوى) مع الأجرعية (الشكل 1).
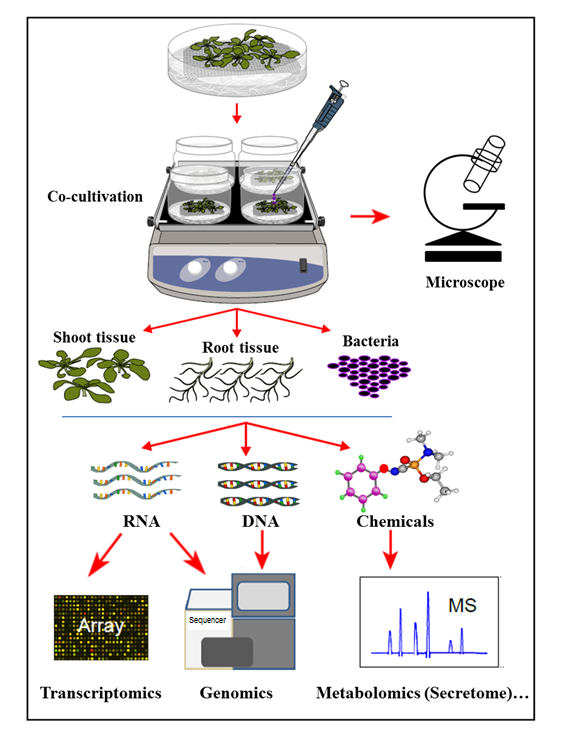
الشكل 1: نظرة عامة على نظام الزراعة المائية، مع تحليلات عينة. تزرع النباتات على رأس شبكة (يطلق النار فوق شبكة)، مع جذور مغمورة في المتوسطة المائية التي يتم بعد ذلك تلقيح مع البكتيريا وأو كوكولتشر. ثم يتم فصل الأنسجة النباتية والبكتيريا للاستخراج في وقت واحد والتحليلات. وقد عدل هذا الرقم من المرجع 35 .
