Repetitive injection of PTZ induces an increase in seizure severity. Six C57BL/6 mice were treated with PTZ, and another 6 mice were treated with saline as a control group. The PTZ dose was 35 mg/kg, and 10 injections were administered. The seizure score gradually increased with PTZ injections, whereas no seizures or abnormal behaviors were evoked by saline injections (Figure 2). ANOVA followed by Bonferroni test showed a significant difference between the PTZ-treated group and the saline-treated group.
Repetitive seizure promotes aberrant axonal branch formation (mossy fiber sprouting) and abnormal migration of granule cells in the hippocampus. Mice were treated with PTZ for 25 injections (dose was adjusted between 24 mg/kg and 35 mg/kg to maintain severe seizure in mice without inducing death caused by a severe seizure). The mouse brains were fixed 3 weeks after the last injection. Control brains were fixed before PTZ injections. Brain slices were immunostained with anti-synaptoporin (x 500) and anti-ZnT3 (x 500) antibodies to observe the mossy fiber sprouting (Figure 3A) and with the anti-doublecortin antibody (x 200) to observe the abnormal migration of granule cells (Figure 3A). Mossy fiber sprouting in the granule cell layer was observed in PTZ-treated slices (Figure 3A). Newborn granule cells, which are immunoreactive for doublecortin, were observed within the hilus in the PTZ-treated slices (Figure 3A). The granule cell layer and hilus shown in Figure 3A is illustrated in Figure 3B.
Repetitive seizures also impair normal behavior of mice. Twelve C57BL/6 mice were treated with PTZ (35 mg/kg, 10 injections), and another 12 mice were treated with saline as a control group. Two weeks after the last injection, the mice were analyzed in a 3-chamber test (Figure 4A) and contextual fear discrimination test (Figure 5A). PTZ-treated mice showed normal sociality (Figure 4B). Mice spent more time in the Stranger 1 chamber than in the Object chamber (saline: p = 0.003, PTZ: p = 0.027) and investigated the Stranger 1 cage more than they investigated the Object cage (saline: p = 0.009, PTZ: p = 0.004). However, PTZ-treated mice showed abnormal social novelty (Figure 4C) indicative of impaired social memory. Control mice spent more time in the Stranger 2 chamber than in the Stranger 1 chamber and investigated the Stranger 2 cage more than they investigated the Stranger 1 cage, whereas the kindled mice did not show any significant difference in time spent in the chambers or time spent investigating the cages (Time in chamber: saline: p = 0.006, PTZ: p = 0.126. Time investigating: saline: p = 0.002, PTZ: p = 0.426). PTZ-treated mice also showed impaired memory in the contextual fear test (Figure 5B). Control mice showed a longer freezing time in the shock condition than in the novel condition, whereas PTZ-treated mice did not show any significant difference in freezing time (saline: p < 0.001, PTZ: p = 0.060). Unpaired t-tests were carried out for statistical analyses.
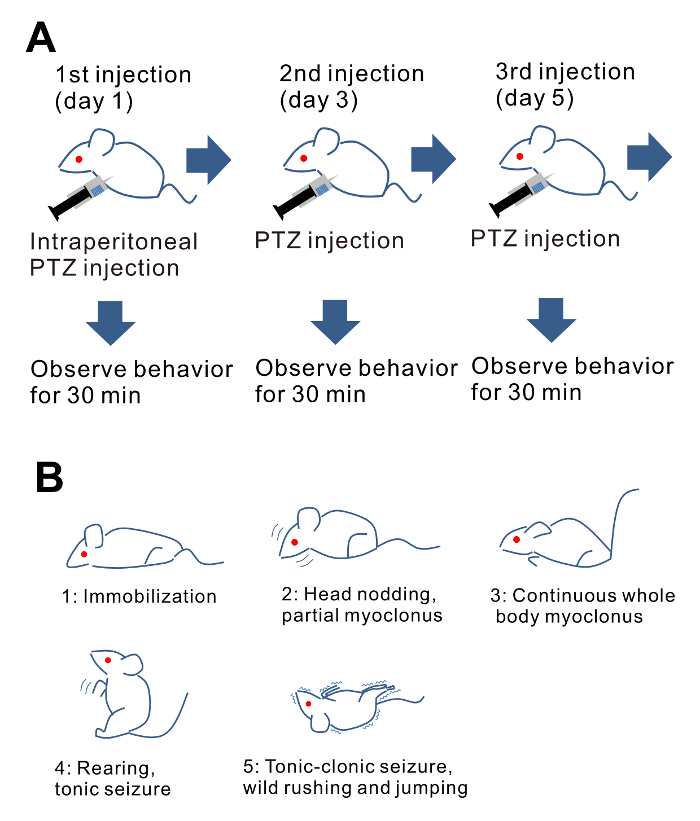
Figure 1: Brief description of the protocol. (A) Schematic illustration of PTZ-mediated kindling. (B) Illustrations of representative animal behaviors for respective seizure scores. Please click here to view a larger version of this figure.
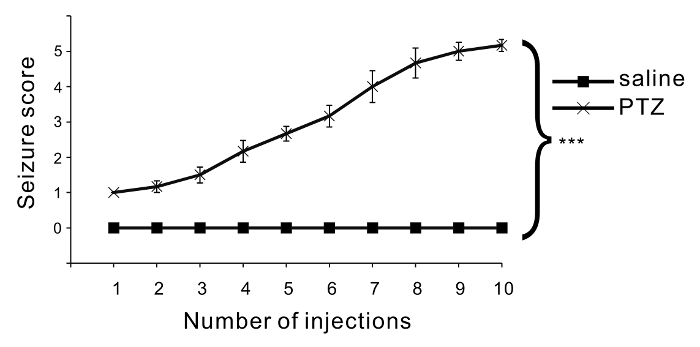
Figure 2: Assessment of convulsive behavior. The mean seizure scores are indicated in the graph. Six mice were used in each condition, and one series of injections was carried out. After each injection, the seizure scores were monitored and scored. Compared to saline injections, PTZ injections significantly increased seizure severity (p < 0.001: repeated-measures ANOVA). Each seizure score is shown as the mean ± SEM. Please click here to view a larger version of this figure.
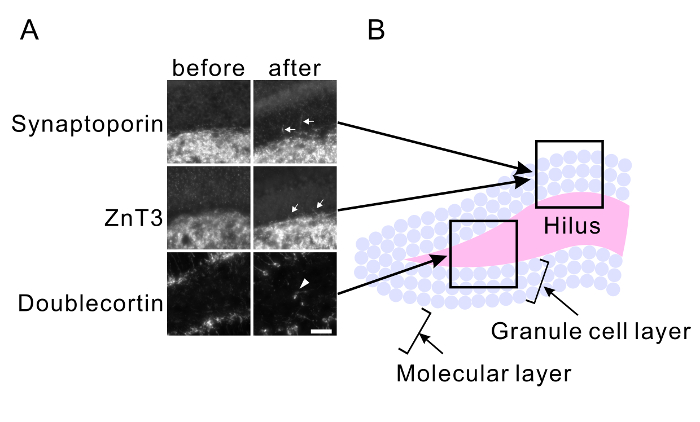
Figure 3: PTZ-mediated mossy fiber sprouting and abnormal migration of granule cells. (A) Maximum projected immunohistochemical images of the hilus, granular layer, and molecular layer of hippocampal slices of the mice before kindling (left) and after kindling (right). Anti-synaptoporin (top) and anti-ZnT3 antibodies (middle) were used to visualize the axons of granule neurons (mossy fibers). The anti-doublecortin antibody was used to visualize newborn granule neurons (bottom). PTZ-mediated repetitive seizures induce mossy fiber sprouting (arrows) and abnormal migration of granule cells into the hilus (arrowheads). Scale bar, 50 μm. The approximate position of each image is indicated in (B). Please click here to view a larger version of this figure.
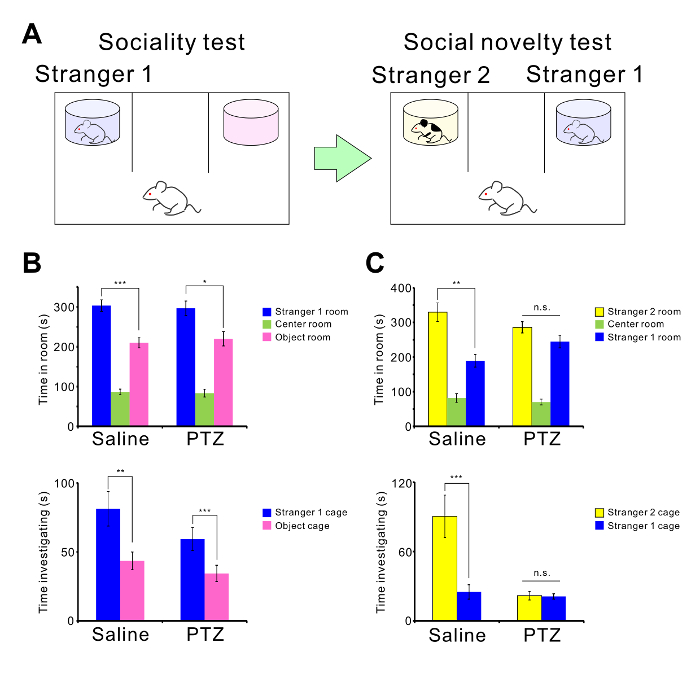
Figure 4: PTZ-mediated abnormal social behavior. (A) Schematic illustration of the 3-chamber test. (B) The mean amount of time spent in each chamber and the mean amount of time spent investigating each cage in the sociality test are shown. (C) The mean amount of time spent in each chamber and the mean amount of time spent investigating each cage in the social novelty test are shown. There were twelve mice in both the PTZ- and saline-treated groups. PTZ-mediated seizures induced abnormal social behavior (***p < 0.001, n.s. = not significant). All times are shown as the mean ± SEM. Please click here to view a larger version of this figure.
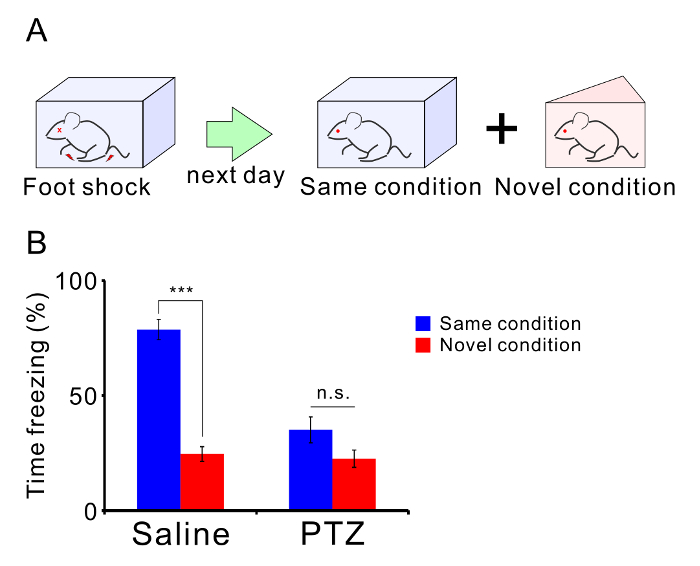
Figure 5: PTZ-mediated memory impairment. (A) Schematic illustration of the contextual fear discrimination test. (B) The mean percentage of freezing times in each condition are shown. There were twelve mice in both the PTZ- and saline-treated groups. Graph shows the mean ± SEM. Please click here to view a larger version of this figure.
