We first sought to investigate whether expressed human breast milk or pasteurized donor breast milk had an effect on small intestinal enteroids. Indeed, human breast milk and donor breast milk increased the growth of neonatal mouse (Figure 1A) and premature human derived enteroids (Figure 1B).
Since human breast milk increased the growth of small intestinal enteroids, we next investigated if breast milk had an effect on the proliferation of premature human enteroids. We demonstrate that human breast milk increases the proliferation of premature human enteroids as compared to control (Figure 2A–B, Ki67, red staining). Importantly, human donor breast milk also increased enteroid proliferation compared to control, but to a slightly lesser degree than non-pasteurized human breast milk (Figure 2B–C). We next evaluated the Ki67 mRNA expression in the premature human enteroids and discovered that Ki67 was increased in the enteroids treated with either breast milk or donor breast milk relative to control (Figure 2D).
We next wanted to see if these effects were species-specific with regards to breast milk. We developed a way to collect breast milk from lactating mice using a modified commercially available human breast pump as in references11,12. Depicted is an anesthetized lactating dam undergoing milk collection using this device (Figure 3). The next series of experiments was performed on mouse enteroids and we evaluated the proliferation effects of expressed breast milk from either mouse, human, or donor breast milk. As shown in Figure 4, mouse, human, and pasteurized donor breast milk increased proliferation in the mouse enteroids as compared to control (Figure 4A–D). Proliferation in the mouse enteroids was decreased in the presence of the TLR4 ligand, lipopolysaccharide (LPS), compared to control (Figure 4A, E). Importantly, mouse, human, or donor breast milk all restored enteroid proliferation in the presence of LPS as demonstrated by increased Ki67 staining compared to LPS alone (Figure 4F–H).
From our experiments, we show that small intestinal enteroids derived from neonatal mice and premature human fetal intestine can be maintained in culture. We further provide a novel breast milk collection method that provides an efficient way to extract breast milk from mice, which can then be used for various experiments. We discovered that expressed breast milk from humans or mice increased the size, growth, and proliferation in mouse and human enteroids. Moreover, mouse, human, and donor breast milk restored the enteroid proliferation after LPS exposure. This suggests that breast milk can exert anti-inflammatory effects on the small intestine of mice and humans.
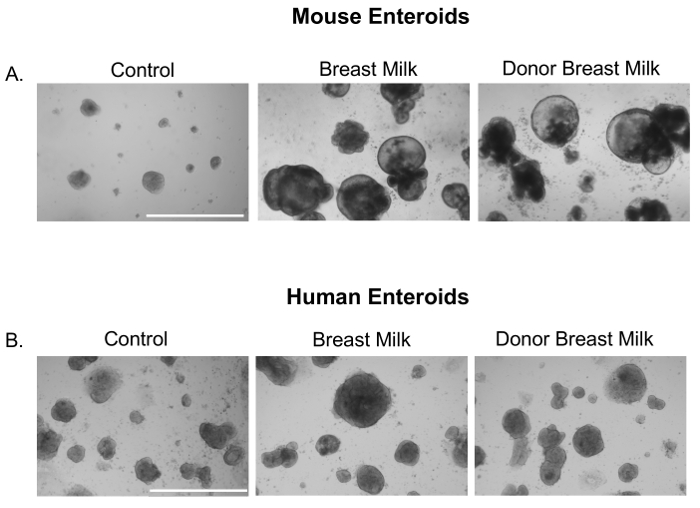
Figure 1. Breast milk and donor breast milk increase the size and growth of mouse and premature human enteroids. Photomicrographs of small intestinal enteroids from neonatal mice (p14) or premature human intestine treated with either media (Control), breast milk, or donor breast milk (50 µL/mL) for 24 h. Size bar: 1,000 µm. Please click here to view a larger version of this figure.
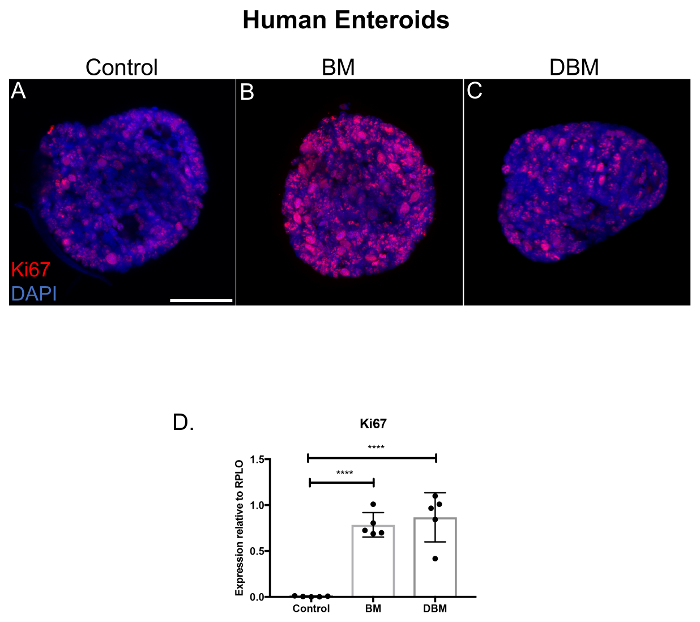
Figure 2. Breast milk and donor breast milk increase the proliferation of premature human enteroids. (A) Confocal micrographs of premature human enteroids cultured for 5 days, treated with human breast milk and pasteurized donor breast milk for 24 h and stained for the proliferation marker Ki67 (red) and DAPI (nuclear, blue). Size bar: 50 µm. (B) qRT-PCR for Ki67 expression of human enteroids relative to the housekeeping gene RPLO. n = 5 per group. **** p <0.0001 comparisons with Student t test compared to control. Data are mean ± standard deviation. Please click here to view a larger version of this figure.
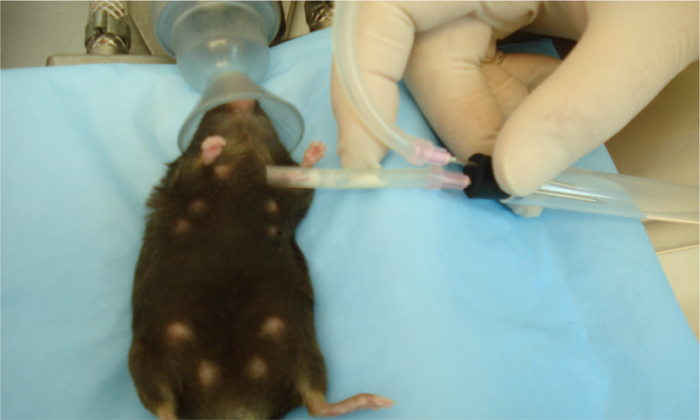
Figure 3. Collection of milk from an anesthetized mouse using a modified human breast pump. Sterile human breast pump tubing placed on teats of anesthetized lactating dam after receiving oxytocin injection. Mouse milk is collected using the human breast pump on medium speed and power into a 5 mL collection tube. Collective volumes extracted from all teats range from 500 µL to 1 mL per mouse at postnatal days 7 – 12. Please click here to view a larger version of this figure.
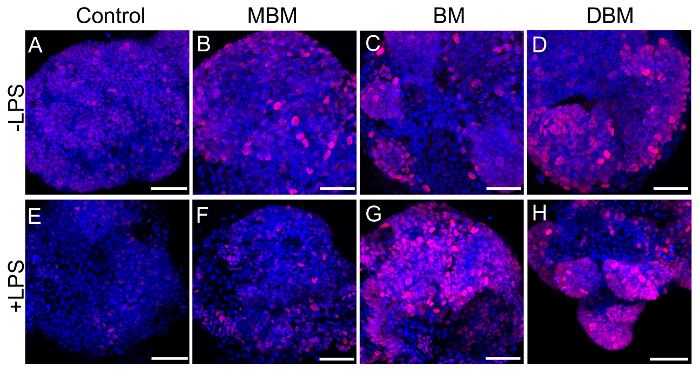
Figure 4. Mouse, human, and pasteurized donor breast milk enhance mouse enteroid proliferation. (A-H) Confocal micrographs of p14 small intestinal mouse enteroids (Control, A) treated with mouse breast milk (MBM, B), human breast milk (BM, C), or human donor breast milk (DBM, D) at 50 µL/mL of media for 12 h in the absence (A-D) or presence of lipopolysaccharide (LPS) at a dose of 25 µg/mL of media for 12 h, (E-H) and stained for the proliferation marker Ki67 (red) and DAPI (nuclear, blue). Size bar: 50 µm. Please click here to view a larger version of this figure.
