Hipoxia, es decir, disuelto las concentraciones de oxígeno [hacer] lo suficientemente bajo para afectar biológica y procesos ecológicos pero a menudo funcionalmente definición como [hacen] < 2 mg / L1y anoxia (funcionalmente definido como [] de 0.0-0.2 mg/L) ocurre más con frecuencia y seriamente en aguas costeras de todo el mundo, estuarios y el océano profundo2,3 y son a menudo exacerbados por aumento de la eutrofización4,5. Con un creciente grado de areal de hipoxia y anoxia, macrofauna se ven afectadas negativamente y perder parte del hábitat y calidad del hábitat. El cambio climático se prevé que empeoran la hipoxia y anoxia6.
En muchos estuarios estratificados, enriquecido con nutrientes tales como Bahía de Chesapeake, Estados Unidos, estacional persistente hipoxia puede prevalecer y puede ocurrir año tras año2. Además, diel-ciclismo de la hipoxia es frecuente en los estuarios como la bahía de Chesapeake y en otros lugares y se produce tarde durante la noche o la madrugada en el verano7,8.
Mayoría de los estudios se ha centrado en el efecto de la continua exposición de organismos a baja [] y en su tolerancia a la hipoxia y anoxia9,10,11,12,13,14 . Por otra parte, estudios han observado el cambio a gran escala en la composición de distribuciones y abundancias de especies las especies en respuesta a la prolongada baja []4,15. A menudo las especies que son muy sensibles a bajas [], mueren en masas,16 cambiando las restantes especies de una fauna menor, menor tamaño, de breve duración como, por ejemplo, encontradas el estante de Louisiana-Texas ecosistema4.
Cambios de comportamiento suelen preceden comunidad colapso17 y estudios han informado sobre las respuestas del comportamiento de los organismos a extendido bajo []4,16,17,18,19 ,20,21,22,23,24,25. Estos estudios, sin embargo, no se centran en las respuestas de los organismos a exposiciones diel-ciclismo de la hipoxia y la naturaleza fluctuante de la disponibilidad [] en los estuarios.
Hipoxia diel-ciclismo en estuarios poco profundos ha recibido cada vez mayor concienciación como estudios de seguimiento [hacer] más frecuentemente en el transcurso de días con sondas en estuarios16,26. Agua puede permanecer hipóxico horas al final de la noche o madrugada en el verano cuando no hay fotosíntesis generadora de oxígeno durante la noche pero el alto consumo de oxígeno respiración aeróbica7,16. También se descubrió que las mareas afectaron el diel ciclismo de condiciones bajas de con la más extrema mínimos observados cuando la marea baja coincidió con el fin de la noche27. Sólo después de varias horas de hipoxia [hacer] volver a normoxia7,16,28 en el ciclo diario.
Para determinar la respuesta conductual de C. virginica y diel-ciclismo hipoxia pH supervisamos la apertura y cierre de las válvulas de las ostras expuestas a laboratorio inducida por diel cíclico y ciclo de [DO] pH. Respuestas de la boca de los bivalvos se han utilizado para detectar condiciones ambientales adversas. Cierre de válvula de bivalvos en respuesta a contaminantes29,30,31, alga tóxica32,33,34, contaminación térmica35,36 , 37, disminuir la cantidad de alimentos38,39,40, alimentación régimen39,41, emersión37,42, fotoperiodo43 , se han medido 44, pH45,46y combinado pH y oxígeno disuelto47 . Técnicas de la boca, por ejemplo, incluyen observaciones directas48,49,13, mediciones continuas usando interruptores reed y los imanes (Dreissena monitor)50o sensores de fibra óptica 51 que requieren agua limpia. Además, se han utilizado sensores de Hall imán y el campo magnético fuerza mejillón gape ángulo52,53,54,55, y un sistema de inducción electromagnética de alta frecuencia que puede medir la distancia variable entre dos bobinas eléctricas que están pegados en las válvulas ha sido utilizado56,57,58,59. Una fuente de alto voltaje se requiere para el sistema de inducción electromagnética y energía tiene a ambos lados de la carcasa52. Este sistema también está disponible comercialmente como el “MOSSELMONITOR” (http://mosselmonitor.nl/).
En un presupuesto apretado investigación, construimos un monitor barato galgas (SGM) para medir continuamente la boca de la ostra sobre diel inducida por laboratorio ciclo de [hacer] y el pH, bajo condiciones de baja visibilidad. Nuestro sistema también es mucho más simple que los sistemas de la competencia, permitiendo a muchos animales a ser instrumentados durante un experimento. Quisimos determinar las respuestas conductuales de C. virginica a diel ciclismo severa ([] = 0,6 mg/L) hipoxia con tampón (pH = 7.8) y ciclismo de pH (pH = 7.8 7.0), respectivamente y boca suave ([] = 1,7 mg/L) hipoxia. Por otra parte, queríamos determinar si las ostras son capaces de responder rápidamente a los cambios en [] sobre el diel ciclo y cómo responder cuando normoxia regresa después de un evento hipóxico. Tal vez las ostras son óptimamente adaptadas al entorno rápidamente fluctuante que se encuentra en muchos estuarios16,27 donde viven. Mientras más complejos monitores gape de válvula están disponibles, el SGM ofrece una técnica barata que permite mediciones continuas de la boca de la válvula en aguas incluso en condiciones de baja visibilidad.
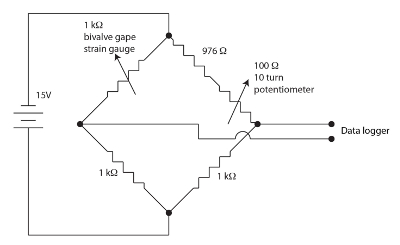
Figura 1. Puente de Wheatstone para el aparato de válvula boca. Haga clic aquí para ver una versión más grande de esta figura.
Los sensores del calibrador de tensión usados para monitorear gape bivalvo son películas resistivas en un patrón del meandro en un polyimide del forro. Pequeñas cantidades de tensión modulan la resistencia del sensor. El bivalvo flexiona las galgas extensiométricas cuando gapes provocando un cambio en la resistencia del sensor. Se empleó un puente de Wheatstone suprimiendo, equilibrado, para cada canal de bivalvo como se muestra en la figura 1 para medir el cambio en la resistencia del sensor. El puente de Wheatstone es nula por el potenciómetro que permite una ganancia bastante alta ser empleado por el datalogger. Un puente de Wheatstone es un método estándar para medir con exactitud una resistencia desconocida usando una proporción de una resistencia conocida estándar y un voltímetro. La historia de esta antiquísima técnica se discute en Ekelof (2001)60. Hemos integrado 12 canales, cada uno con su propio puente de Wheatstone y suprimiendo el potenciómetro, en la unidad de la galga de tensión Monitor (SGM).
