Oligodendrocyter (OLs) är myelinating celler i det centrala nervsystem (CNS)1. Isolering och kultur av primära oligodendrocyter i en hårt reglerad miljö är ett värdefullt verktyg för in vitro- studier av utvecklingen av oligodendroglia samt biologi demyeliniserande sjukdomar såsom multipel skleros2 . Detta kräver en effektiv och robust oligodendrocyte isolering och kultur metod3. I denna studie tog vi fördel av uttrycket av en distinkt oligodendrocyte cell surface markör att genomföra en modifierad isolering teknik som är snabb och specifika.
Fyra olika faser av oligodendrocyte mognad har identifierats, var och en kännetecknas av uttrycket av särskiljande cell yta markörer för varje utvecklingsfas (figur 1). Dessa cell yta markörer kan kännas igen av specifika antikroppar4,5, och kan användas för att isolera OLs i specifika stadier. I den första etappen har oligodendrocyte föregångare celler (OPCs) förmåga att föröka sig, migrera och specifikt express trombocyt-härrör tillväxtfaktor receptor (PDGF-Rα)6, ganglioside A2B5, proteoglykan NG27,8 , polysialic syra-neurala cell adhesion molekyl9 och fettsyror-syra-bindande protein 7 (FABP7)10. OPCs har bipolär morfologi med några korta processer som härrör från de motsatta polerna av cell kroppen, vilket är karaktäristiskt för neurala föregångare celler11.
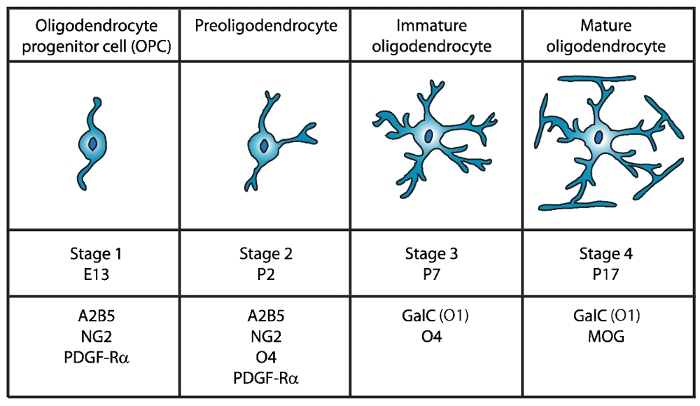
Figur 1: uttryck för cell yta markörer under musen oligodendrocyte utvecklingen. OLs cell yta markörer såsom A2B5, GalC (O1), NG2, O4 och PDGF-Rα kan användas för att isolera specifikt oligodendrocyter på särskilda utvecklingsstadier med hjälp av specifika antikroppar. Vänligen klicka här för att visa en större version av denna siffra.
I den andra etappen, OPCs ge upphov till preoligodendrocytes och express på cellmembranet inte bara OPC markörer, men också den sulfatide (en sulfaterade galactolipid) erkänns den O4 antikropp12,13och GPR17 protein14 som framhärdar till omogna oligodendrocyte (OL) scenen. I detta skede förlänga preoligodendrocytes multipolär kort processer. Preoligodendrocytes är den stora OL-scenen på postnatal dag 2 (P2) i den cerebrala vita substansen i både råtta och mus med en mindre befolkning av omogna OLs15.
Under den tredje etappen fortsätta omogna OLs att uttrycka O4, förlora uttryck för A2B5 och NG2 markörer och börjar uttrycka galactocerebroside C16. I detta skede OLs är engagerade av oligodendroglial härstamning och bli efter mitotiska celler med långa förgrenat grenar17,18. Omogna OL utgör mer än 80% av de gnagare vit substansen på P7 och vid denna tid de första MBP+ -cellerna observeras15,19,20,21. Isolering av OLs på P7 kunde därför säkerställa cellernas kickavkastning.
I det sista och fjärde utvecklingsstadiet OL express mogen OLs myelinating proteiner (myelin grundläggande protein (MBP), proteolipid protein (PLP), myelinet förknippas glykoprotein (MAG) och myelin oligodendrocyte glykoprotein (MOG)22,23 ,24,25,26. I detta skede mogen OLs förlänga membran att formuläret kompakt omsluter slidor runt axoner och kunna myelinate. Detta sammanfaller med observationen att i råtta och mus hjärna, MBP+ celler blir alltmer riklig P1419,20,21.
Sedan den första isoleringen av oligodendrocyte av Fewster och kollegor i 196727, har flera metoder för isolering av OLs från gnagare CNS genomförts inklusive immunopanning28,29,30, fluorescens-aktiverad cell sortering (FACS) att utnyttja cell surface-specifika antigener28,31, differentiell gradient centrifugering32,33,34,35 och en skakning metod baserad på differentiell följsamhet av olika CNS glia36,37. Befintliga kultur metoder har dock begränsningar, särskilt när det gäller renhet, avkastning och tid som krävs för att utföra de förfarande38. Därför krävs effektivare isoleringsmetoder för oligodendrocyter.
I detta papper, vi presentera ett enkelt och effektivt urvalsmetod för immunmagnetisk isolering av etapp tre O4+ preoligodendrocytes celler från neonatala möss pups. Denna metod är en modifiering av de tekniker som rapporterats av Emery et al. 39 och Dincman o.a. 40 och ger en oligodendrocyte förberedelse renhet över 80% i ca 4 h.
