Cardiomyocytes (CMs) dérivés des cellules souches pluripotentes induites (CISP) sont un outil émergent à décortiquer les mécanismes moléculaires des maladies cardiaques, d’enquêter sur nouveaux traitements et à l’écran pour cardiaques indésirables effets1,2 ,3. Dès le début, arythmogène maladies tels que les canalopathies ont été un objectif important de ce domaine de recherche4. Par conséquent, les méthodes pour étudier des phénotypes électriques de CMs, comme arythmies ou des changements dans la morphologie du potentiel d’action (PA), sont au cœur de cette technologie.
Une considération importante dans l’application de l’iPSC-CMs est que les protocoles actuels de différenciation cardiaques n’entraînent pas une population homogène de cellules. Au lieu de cela, ils sont plutôt un mélange de cellules ressemblant à n ud sinusal, auriculaire et ventriculaires CMs à différents niveaux de maturation5,6,7,8. Cette hétérogénéité peut être une source pertinente de la variabilité expérimentale, surtout si les paramètres tels que la durée des PA (APD) sont l’objet d’une enquête, qui diffèrent intrinsèquement entre sous-types CM (p. ex., l’APD est plus courte dans auriculaire que dans CMs ventriculaires). L’approche classique pour régler ce problème est d’enquêter sur iPSC-CMs unique à l’aide de la méthode de serrage de patch et de classer chaque cellule sous la forme nodale-, auriculaire-, ou ventriculaire-like, basée sur la morphologie de son AP9. Toute analyse ultérieure peut être restreint alors aux cellules représentant le sous-type de CM d’intérêt. L’inconvénient majeur de cette stratégie est son débit limité et le manque d’évolutivité. En outre, le caractère invasif de l’électrophysiologie de patch clamp ne permet pas de l’imagerie des cellules mêmes séquentiellement sur une période de temps prolongée.
Ici, nous fournissons des détails expérimentaux sur une méthode10 mis au point pour l’image optiquement APs en sous-types spécifiques de l’iPSC-CMs. Cela permet de surmonter le problème de l’hétérogénéité de sous-type et augmente considérablement le débit par rapport aux méthodes conventionnelles, permettant le phénotypage rapide du CISP-CMs transportant des variants génétiques ou d’être des agents exposés à pharmacologique.
Aperçu de l’approche d’imagerie optique du sous-type spécifique
Un indicateur de tension codé génétiquement (GEVI), dont les propriétés fluorescence changent lors de la dépolarisation et la repolarisation de la membrane cellulaire, est utilisé pour image optiquement les changements du potentiel membranaire des CMs. Le GEVI appliqué ici est la protéine fluorescente-détection de tension VSFP-CR11, qui se compose d’un domaine transmembranaire détection tension fusionné à une paire d’un vert (trèfle) et une protéine fluorescente rouge (mRuby2) (Figure 1 a). En raison de la proximité des deux fluorophores, l’excitation de la protéine fluorescente verte résulte en une fraction de l’énergie d’excitation étant transféré à la protéine fluorescente rouge par transfert d’énergie de résonance Förster (FRET). Par conséquent, l’excitation de la protéine fluorescente verte entraîne une émission à la fois le vert et les rouges protéines fluorescentes (Figure 1 a, panneau supérieur). Lorsque la cellule dépolarise, un réarrangement structural de la sonde de tension se produit qui se traduit par une réorientation des deux protéines fluorescentes, augmentant ainsi l’efficacité de la frette. Ainsi, encore plus de l’énergie d’excitation est transféré du vert à la protéine fluorescente rouge (Figure 1 a, panneau inférieur). Ainsi, dans une cellule dépolarisée, l’émission de fluorescence verte est plus faible, et l’émission de fluorescence rouge est plus brillante que dans une cellule au repos le potentiel de membrane (Figure 1 b).
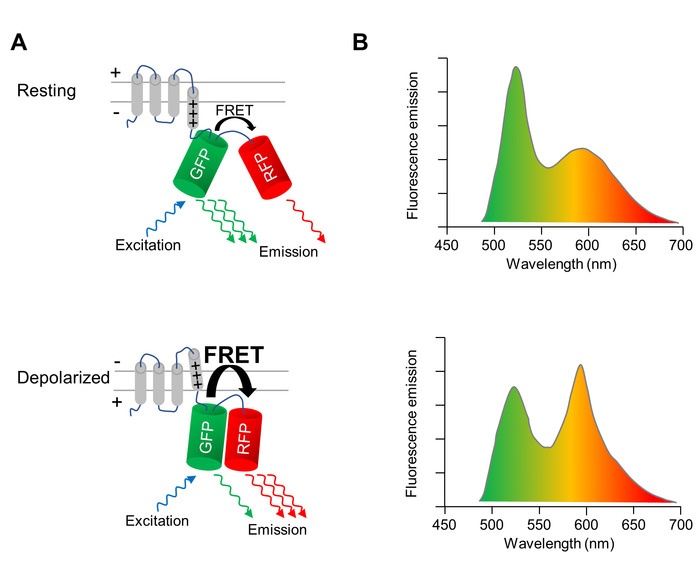
Figure 1 : l’imagerie optique du potentiel avec VSFP-CR. de membrane (A) A schéma illustrant l’action de la protéine fluorescente sensibles à la tension QUE VSFP-CR est montré. Lors de la dépolarisation de la membrane cellulaire, un réarrangement structural du domaine transmembranaire-détection de tension se traduit par une réorientation de la verte (GFP) et rouge protéine fluorescente (DP), augmentant l’efficacité de la Förster intramoléculaire transfert d’énergie par résonance (FRET). Les spectres (B) l’émission d’une VSFP sur l’excitation de la GFP dans les cellules au potentiel de repos membranaire (panneau du haut) et dans les cellules dépolarisées (panneau inférieur) sont représentés. Le changement spectral sur dépolarisation est exagéré pour plus de clarté. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
Les changements dans l’efficacité de la frette reflétant les fluctuations du potentiel membranaire sont imagés à l’aide d’un microscope à fluorescence équipé d’un séparateur de l’image, qui sépare l’émission de fluorescence rouge et vert et les projette sur deux zones adjacentes de la puce d’une caméra sCMOS (Figure 2). Avec ce set-up, l’émission de fluorescence à deux bandes de longueur d’onde différente peut être enregistrée simultanément, ce qui permet le calcul d’un ratio de fluorescence rouge au vert afin de refléter le potentiel de membrane dans chaque image d’une série de Time-lapse.
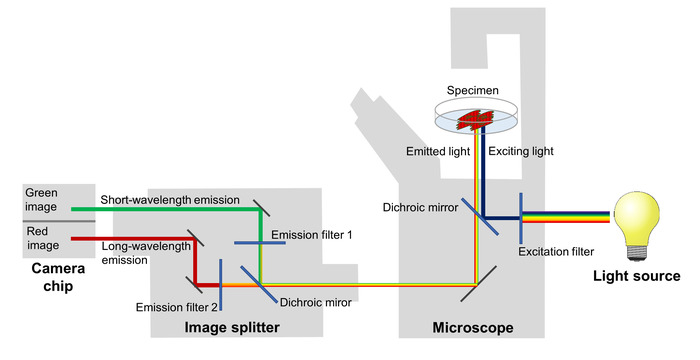
Figure 2 : Configuration du système d’imagerie. Les principaux composants du système d’imagerie utilisé pour image les modifications spectrales de la protéine fluorescente sensibles au voltage, reflétant les changements potentiels de membrane à une haute résolution temporelle sont représentées. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
L’expression de VSFP-CR dans CMs est obtenue par transduction des gènes. Pour diriger l’expression au sous-type CM d’intérêt, le lentivirus contient un élément de promoteur (le renforceur de MLC2v ) qui anime spécifiquement transcription ventriculaire type iPSC-CMs10. Lorsque l’iPSC-CMs qui représentent un mélange de cellules de type auriculaire, nodal, ventriculaire-type et sont transduites avec ce lentivirus, VSFP-CR est exprimée uniquement dans les cellules ventriculaires. Étant donné que l’imagerie optique potentiel d’action dépend de cette sonde fluorescente, les potentiels d’action enregistrés représentent exclusivement le sous-type de CM d’intérêt (Figure 3).
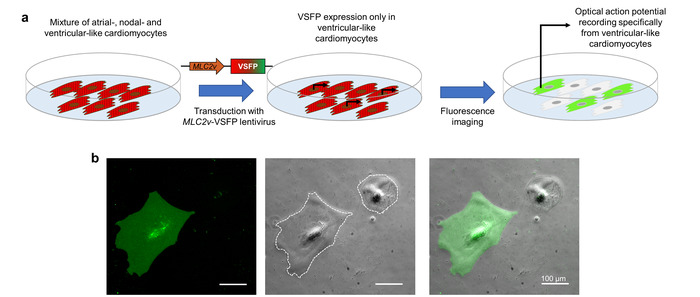
Figure 3 : expression VSFP pilotée par le promoteur pour l’imagerie de potentiel membranaire sous-type spécifique. (a) ce schéma montre comment enregistrements de potentiel d’action optique sous-type spécifique cardiomyocyte sont atteints. (b) iPSC-CMs infectés par un VSFP sous le contrôle de le MLC2v-amplificateur de ventriculaire spécifiques sont indiqués. L’expression de la sonde de tension est observée seulement dans CMs ventriculaire-comme dans le chenal de la GFP (panneau de gauche). Le contraste de phase (panneau central) et une image de superposition (panneau de droite) sont également fournis. Les pointillés blancs marquent les limites d’une cellule. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
