חלבון אינטראקציות אני חוץ גופית n
באופן מסורתי, קריסטלוגרפיה וניסויים תהודה מגנטית גרעינית בשילוב עם מיקרוסקופ אלקטרונים הקפאה (cryoEM) הם הטכנולוגיות שבחרת לתאר במדויק את הארכיטקטורה תלת-ממדי של חלבונים, להסיק שלהם פונקציה על ידי המתחקה אחר פרטיהם מבניים ברזולוציה גבוהה. חלבונים, לעומת זאת, אינם מבנים סטטי ולא ניתן לעבור מגוון של התנודות והשינויים הסתגלותי בזמן ובמרחב. זו הסיבה מדוע מבניים מידע לחקר הגבישים או נתונים CryoEM צריך השלמה עם טכניקות אחרות (למשל, דינמיקה מולקולרית סימולציות וטכניקות מולקולה בודדת): הפונקציה של חלבון קשורה שלה הסתגלותי שינויים אינטראקציות, ואת המידע הזה אינו נוכח בתוך מבנה סטטי. על מנת לחקור דינמיקה מולקולרית התוך, טכניקות בהתבסס על מולקולה בודדת פורסטר תהודה להעביר אנרגיה (smFRET) הן יעיל מאוד1. גישות אלה מסוגלים להעריך subpopulations שונים של מולקולות מורכבות בתקשורת. זה חשוב מאוד, כמו שינויים אלה מהירים להתרחש במהלך רכישת הנתונים (קרי, הננו לטווח השני).
שתי גישות עיקריות מועסקים בדרך כלל כדי לזהות ולכמת שינויים אלה: חלבונים פתרון ואת השטח-הנייח. איתור של האינטראקציות הבין-מולקולריות ובמיוחד, התהליך של dimerization המושרה על ידי ליגנדים, smFRET אינה תמיד הכלי הטוב ביותר. אכן, סריג תלוי לא רק על המרחק (≈10 ננומטר), אלא גם על הכיוון של שני הדיפולים (התורם ולא מקבל, χ2) ונמצא החפיפה של הפליטה תורם עם ספקטרום הבליעה של מקבל2, אבל אולי המצב האחרון הזה חשוב ובלבד experimentalist יכולה בחרה הזוג סריג הנכון. חיסרון מסוים של smFRET דורשים בדיקה ההומו-dimerization נובע תיוג של החלבון עניין: עבור הטרו smFRET, dimerization ניתן רק זוהה עד 50% (קרי, הטרו-סריג יהיו רק יכול לזהות את התורם-מקבל, התורם מקבל הומו-הדימרים אך לא תורם-תורם או מקבל-מקבל, המהווה השני 50% הדימרים). השימוש של קרינה פלואורסצנטית המתאם ספקטרוסקופיה (FCS) נגזרים (FCCS, ועוד3) כדי לברר חלבון דיפוזיה קבועים, איגוד קבועים במבחנה הוא חלופה אחרת. גישות אלה אינם מסוגלים לחלוטין לכמת ההומו-dimerization גם, כמו FCS אחד האמצעים מקדם ריכוז, דיפוזיה, ואת רדיוס דיפוזיה של חלקיק באמצעי תלויים בצורה לא טובה המשקל המולקולרי; עלייה 10-fold משקל מולקולרי רק לרמוז לדוגמה שינוי 2.15 לקפל מקדם דיפוזיה4. במקרה של FCS שני צבעים או FCCS, רק 50% של הומו-הדימרים יראו מאותה סיבה כמפורט לעיל. גישות מעשיות ביותר כמותיים כדי לזהות הומו-dimerization במבחנה וב -vivo הם הומו-סריג5 , מספר, בהירות (N & B)6. בהתחשב בעובדה הומו-סריג דורש שחזור במכשור ספציפי של הערך חיזקו (קרי, אופטי/אנלייזרים יסודות לשחזר את קיטוב מקבילים, בניצב), N & B מוצג כאן כמו טכניקה חיובית מזהים חלבון ההומו-dimerization, צבירה. זה יכול להיות מועסק גם במבחנה וגם ויוו עם תחבולה מסחרית.
מספר ובהירות
N & B כבר שנסקרה לאחרונה7. הביקורת התמקדה ביישום הטכניקה תאים חיים. זה משתלם כדי לשכפל פורמליזם מתמטי כאן משוואות אלה יוחלו על הנתונים שנאספו בתוך חוץ גופית. ראשית, יש צורך להגדיר כמה מונחים וכמויות מתמטי:
- ישות היא קבוצת מולקולות אשר קשורים אחד לשני.
- חדוה הבהירות של ישות הוא המספר של פוטונים שהוא פולט ליחידת זמן (לכל מסגרת).
- n הוא המספר של ישויות הנוכחי.
- של פיקסל נתון במהלך סדרות התמונה, הוא שלה כלומר עוצמת ו σ2 הוא השונות בעוצמתה.
לאחר מכן, עם ספירת פוטון גלאי, בהנחה ישויות ניידים, ללא רקע,
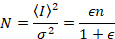
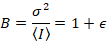
כאשר N הוא מספר לכאורה ו- B זה בהירות נראית לעין. התוצאה היא
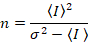
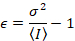
דלאל. et al. 8 הראה כי עם ציוד אנלוגי, אחד צריך תיקון 3 תנאים: על גורם S, על רקע היסט, רעש readout σ02. לאחר מכן, שוב בהנחה ישויות ניידים,
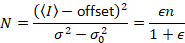
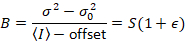
נותן
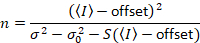
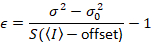
שימו לב כי המשוואה הנ עבור שונה זה נתון דלאל. et al. 8 וסקירה עוקבות. 7 ב דלאל. ואח ה- S במכנה הושמט בשל טעות דפוס, שגיאה זו שוכפלה על הסקירה. המשוואה הנ ל היא הנכונה. הנחיות למדידת S, σ והיסט02 – יחד עם הסבר על משמעותם – ניתנים על-ידי דלאל. et al. 8
חדוה הבהירות הוא יחסי המדינה oligomeric של הישויות באמצעי: חדוה יהיה פעמיים גדול עבור הדימרים כמו זה מונומרים, גדולה פי שלוש עבור trimers כפי שהיא מונומרים, פעמיים גדול עבור hexamers כפי שהוא trimers וכן הלאה. בדרך זו, מודדים את הבהירות חדוה, אחד יכול לכמת כל סוג של multimerization.
אם יש תערובת של oligomeric הברית הנוכחי, מספר והבהירות אינה מסוגלת להתאושש הברית oligomeric בודדים הנוכחי. זוהי מגבלה של הטכניקה.
Detrend אלגוריתם ותוכנות nandb
החשיבות של תיקון עבור photobleaching כבר הדגיש בעבר9. Photobleaching באופן בלתי נמנע מתרחשת במהלך הניסויים מיקרוסקופ אור במצב זמן לשגות; שניהם תאים חיים, חוץ גופית בתוך. גישות רבות תוארו בספרות לתיקון הלבנת7. טכניקת סינון מעריכית עם בחירה אוטומטית של פרמטר detrending T הוא הטוב ביותר הנוכחי. היא משולבת לתוך nandb תוכנה חופשית, קוד פתוח9. אכן, תוכנה הדורשים מהמשתמש לבחור באופן ידני את הפרמטר detrending שלהם יכול לגרום תוצאות שגויות בגלל בחירה פרמטר זו צפויה להיות שרירותי של שגוי. האלגוריתם אוטומטיים בודק את הנתונים וקובע את הפרמטר המתאים, ללא צורך התערבות המשתמש9. אפילו עם הבחירה הטובה ביותר של פרמטר המחליק, detrending יש מגבלות משלו ועובד היטב רק עם photobleaching אחוזים נמוכים יותר 25%, כפי שמוצג עם סימולציות9. מעניין, בעת שימוש השגרה detrending אוטומטי, רמת הדיוק שלה היא כזאת אחד יכול לעבוד עם ערכי בהירות נמוכה (אפילו B < 1.01), ומכאן עוצמות, והנמוכות עדיין להיות מדויק מספיק כדי לכמת את ההומו-dimerization.
Photobleaching גורם גם בעיה נוספת: הנוכחות של photobleached fluorophores ב multimer מורכבים. זה עושה למשל, trimer נראות דיימר כאשר אחד של שלוש יחידות ב trimer הלא-פלורסנט. עובד, מולר10 הראה כיצד לתקן בשביל זה, תיקון זה הודגש גם ב- סקירה עוקבות7. התוכנה nandb כולל תיקון זה9.
FKBP12 F36V מערכת
FKBP12F36V הוא חלבון אשר לא oligomerize באופן טבעי אך ידוע dimerize עם התוספת של AP20187 סמים (המכונה בפיהם BB dimerizing ליגנד)11,12. זה עושה את זה מקרה מבחן אידיאלי עבור מספר והבהירות: עם FKBP12F36V עם תוויות ברורות, הכפלה של מדינת oligomeric צריך להקפיד על תוספת של BB.
