תהליכים ביולוגיים רבים להתרחש בכל האתרים של תא-תא אינטראקציות, למשל, תא-תא אדהזיה1,2,3, תא-cell fusion4 וזיהוי הסלולר5. אירועים כאלה חשובים במיוחד במהלך הפיתוח של אורגניזמים רב תאיים, תקשורת, למשל, במהלך תגובות מערכת החיסון. תהליכים אלה הם בדרך כלל בתיווך חלבונים אשר מותאמים על פני השטח, קרי, ב קרום פלזמה (PM) של תאים שכנים ומבצעת אינטראקציות ספציפיות-הקשר תא-תא כי הם בדיוק הזמן והמרחב מוסדר. במקרים רבים, אינטראקציות אלה ישירה הומו – או אינטראקציות חלבון heterotypic טרנס , אך עשוי גם לכלול יונים או ליגנדים מתנהג כמו linkers חוץ-תאי1. למרות חשיבות היסוד, יש מחסור של מבחני הבדיקה אינטראקציות חלבון-חלבון ספציפי אלה ישירות בסביבה הטבעית של תאים חיים. שיטות רבות דורשות גם לשבירת תאים (למשל, הביוכימי מבחני כגון co-immunoprecipitation6), קיבעון (למשל, כמה טכניקות במיקרוסקופ אופטי סופר רזולוציה ו מיקרוסקופ אלקטרונים של תא-תא קשר7), או שאינם ספציפיים, כגון: צבירת / מבחני אדהזיה8,9. כדי להתגבר על בעיה זו, טכניקות קרינה פלואורסצנטית יושמו מבוסס על קרינה פלואורסצנטית תהודה אנרגיה העברה (סריג)10 או קרינה פלואורסצנטית קומפלמנטציה11. עם זאת, כדי להשיג מספיק קטן המרחקים בין fluorophores, שיטות אלה דורשים תוויות פלורסנט בצד חוץ-תאי של חלבונים10, פוטנציאל מתנגש עם טרנס אינטראקציות.
כאן, אנו מציגים חלופי המבוסס על ידי קרינה פלואורסצנטית assay עבור אינטראקציות חלבון-חלבון תא-תא באנשי קשר. גישה זו משלבת גישות קרוס-המתאם קרינה פלואורסצנטית (סריקה קרינה פלואורסצנטית קרוס-המתאם ספקטרוסקופיה (sFCCS), מספר קרוס-המתאם ואת הבהירות (ccN & B)), ערבוב של תאים המבטאים בונה פיוז’ן של החלבון של . עניין, למשל, קולטן אדהזיה. קולטני ובדוקים בתאים שמעצבת שני מסומנות עם שני חלבונים פלורסנט spectrally מופרדים (FPs), מתאיים הצד (ראה איור 1 א’).
השיטות מועסקים מבוססים על ניתוח סטטיסטי של תנודות פלורסצנטיות המושרה על ידי התנועה דיפוזיה של פיוז’ן פלורסנט חלבונים דרך נפח מוקד של לייזר קונפוקלי מיקרוסקופ סורק. יותר בפירוט, וזמינותו רגשים פעפוע שיתוף של החלבונים עניין במחזור שני שכנים תא-תא באנשי קשר. אם החלבונים עוברים אינטראקציות טרנס , אלה מתחמי טרנס ויישא חלבונים פלורוסנט פולטות בשני ערוצים ספקטרלי, גורם תנודות פלורסצנטיות מתואם של שנינו פולטים. מצד שני, אם הכריכה לא מתרחשת, מספר תנודות של חלבונים מול PMs תהיה עצמאית, גורם תנודות לא מתואם. הרכישה יכול להתבצע בשתי דרכים: 1) sFCCS מבוסס על סריקה בצורת קו לאורך הקשר תא-תא, רגשים ביעילות את האינטראקציות במקום הממוקמים באזור הקשר. דרך ניתוח הטמפורלי של תנודות פלורסצנטיות, sFCCS מספקת גם מידע dynamics, קרי, מקדמי דיפוזיה של מתחמי חלבון; 2) ccN & B מבוססת על ניתוח pixel-wise של רצף של תמונות רכשה את האזורים קשר תא-תא. יש יכולת בדיקה של אינטראקציות מפה לאורך כל קשר עם איזור (ב מישור מוקד אחד), אך אינו מספק מידע על דינמיקה. שתי השיטות יכול להיות משולב עם ניתוח של הבהירות מולקולרית, קרי, האות הממוצע פלורסצנטיות הנפלטת ביחידת זמן אחת מתחמי חלבון באמצעי ולספק, לפיכך, הערכות של סטויכיומטריה של מתחמי חלבון- תא-תא אנשי קשר.
במאמר זה, אנו מספקים נתונים היסטוריים פרוטוקולים עבור הכנת הדוגמא, כיול המכשיר, רכישת נתונים וניתוח לבצע את הבדיקה הציג על לייזר קונפוקלי מסחרי מיקרוסקופ סורק. ניתן לבצע את הניסויים על כל כלי מצויד פוטון לספור או גלאי אנלוגי ולא אובייקטיבית עם מפתח נומרי גבוהה. אנו עוד יותר לדבר על שלבים קריטיים של הפרוטוקול, לספק ערכות תיקון עבור מספר תהליכי גורם תנודות אות artefactual, למשל, גלאי רעש, תנועה photobleaching או תא. פותח במקור כדי לחקור את האינטראקציות בין תאים חסיד, וזמינותו עשוי להיות שונה עבור השעיה תאים, או הותאם מודל ממברנה מערכות, למשל, unilamellar ענק שלפוחית (כשמרוגזים) או פלזמה ענקית קרום שלפוחית (GPMVs), המאפשר כימות של אינטראקציות בסביבות השומנים שונה או בהיעדר שלד התא מאורגן12,13.
סריקת קרינה פלואורסצנטית קרוס-המתאם ספקטרוסקופיה גירסה שונה של קרינה פלואורסצנטית קרוס-המתאם ספקטרוסקופיה14 , תוכננה במיוחד כדי לחקור דינמיקה דיפוזיה איטית השומנים ממברנות15. הוא מבוסס על רכישה סריקה קו בניצב רה מ המכיל את החלבונים פלואורסצנט עניין. כדי לחקור אינטראקציות של שני המינים חלבון שונה שכותרתו, הרכישה מתבצעת בשני ערוצים ספקטרלי באמצעות שני קווי לייזר, שני חלונות איתור עבור fluorophores spectrally מופרדים. בשל הדינמיקה דיפוזיה איטית של חלבונים ברה מ (D≤ ~ 1 /s2מיקרומטר), מידה קרוס-לדבר נטול יכול להתבצע על ידי לסירוגין את ערכת עירור משורת בשורה15. הניתוח מתחיל עם: 1) תיקון אלגוריתם יישור לתנועה לרוחב התא מבוסס על block-wise בממוצע של ~ 1000 שורות, 2) קביעת המיקום עם פלורסצנטיות המרביכלומר הממשלה מיקום האות, בכל בלוק, 3) הסטה של כל הבלוקים על נפוצות מקור12,15, בנפרד בכל אחד מהערוצים. לאחר מכן, מבחר אוטומטית של פיקסלים רה מ מתבצע על-ידי בחירה באזור המרכז התאמה לפי עקומת גאוס הסיכום של כל הקווים מיושר (קרי, מרכז ± 2.5σ). שילוב של האותות בכל שורה התשואות ממברנה פלורסצנטיות סדרת הזמן F(t) בכל אחד מהערוצים (g = ירוק ערוץ, r = הערוץ האדום). שים לב כי גודל פיקסל חייב להיות קטן מספיק, למשל, < 200 ננומטר, לשחזר את צורת הנקודה להפיץ את הפונקציה ולמצוא את המרכז שלו, המתאים למיקום של ראש הממשלה. בנוכחות photobleaching משמעותית, סדרת הזמן פלורסצנטיות בכל אחד מהערוצים ייתכן המודל עם פונקציה מעריכית-כפול ויש לאחר מכן תוקן עם הנוסחה הבאה:16
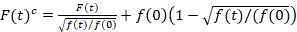 . (1)
. (1)
חשוב לציין כי נוסחה זו מתקנת ביעילות amplitudes והן דיפוזיה פעמים המתקבל ניתוח המתאם של F(t)c, לעומת הערכות פרמטר זה ניתן להשיג את שלא תוקנו F(t). ואז, הפונקציות אוטומטי – ואת הצלב-קורלציה (ACFs / CCFs) של קרינה פלואורסצנטית החישוב של אותות:
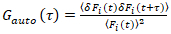 , (2).
, (2).
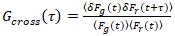 , (3).
, (3).
איפה אלפאFאני = Fאני(t) – 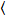 Fאני(t)
Fאני(t)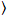 , ואני = g, r.
, ואני = g, r.
מודל דו מימדי דיפוזיה מצויד ואז המתאם לכל הפונקציות (CFs):
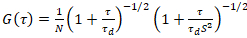 . (4)
. (4)
כאן, N מציין את מספר חלבונים פלורסנט אמצעי תצפית, τd הזמן דיפוזיה לכל ערוץ. מודל זה לוקח בחשבון כי בהגדרה ניסיוני המתואר, דיפוזיה של חלבונים ברה מ מתרחשת בתוך המטוס x-z, בניגוד נפוץ התצורה של המתאם פלורסצנטיות ספקטרוסקופיה (FCS) ניסויים ממברנות חיטוט דיפוזיה במישור x-y של נפח קונאפוקלית17. המותניים w0 והגורם מבנה S, המתארת את התארכות wz של אמצעי האחסון נקודתית ב- z, S = wz/w0, מתקבלים מ כיול נקודת FCS מדידה המבוצעת עם spectrally דומים צבעי והגדרות באותו האופטי באמצעות הערכים הזמינים כבר מקדם דיפוזיה Dצבע:
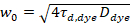 , (5).
, (5).
איפה τd, צבע זה הזמן נמדד פיזור ממוצע של מולקולות צבען, המתקבל מתאים למודל תלת מימדי דיפוזיה בנתונים, לוקח לתוך חשבון מעברים של השבר T של כל המולקולות N שלישיה המדינה עם קבוע הזמן ττ:
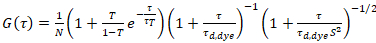 . (6)
. (6)
לבסוף, דיפוזיה (D), ערכי הבהירות מולקולרית (חדוה) והקבוע יחסית קרוס-הקורלציה של נתונים sFCCS (rel.cc.) מחושבים כדלהלן:
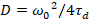 , (7).
, (7).
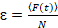 , (8).
, (8).
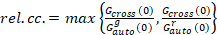 , (9).
, (9).
Gקרוס(0) איפה משרעת הפונקציה קרוס-קורלציה, 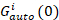 הוא משרעת הפונקציה autocorrelation בערוץ אני-th.
הוא משרעת הפונקציה autocorrelation בערוץ אני-th.
הגדרה זו של יחסי קרוס-המתאם, קרי שימוש מרבי במקום כלומר המשוואה 9, לוקח בחשבון את המספר המרבי של קומפלקסים של חלבונים שני מינים נוכח בריכוזים שונים הוא מוגבל על ידי מינים נוכח מספר נמוך יותר.
בהירות ומספר צלב-המתאם מבוסס על ניתוח רגע של עוצמת קרינה פלואורסצנטית לכל פיקסל של אוסף תמונות רכשה במשך הזמן במיקום קבוע במדגם, המורכב בדרך כלל ~ 100-200 מסגרות, עם שני ספקטרלי ערוצי ( g = ירוק ערוץ, r = הערוץ האדום). מהממוצע טמפורלית אני, ואני השונות 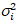 , הבהירות מולקולרית חדוהi ואת מספר nאני מחושבים ב כל פיקסל וערוץ ספקטרלי (אני = g, r)18:
, הבהירות מולקולרית חדוהi ואת מספר nאני מחושבים ב כל פיקסל וערוץ ספקטרלי (אני = g, r)18:
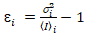 , (10).
, (10).
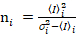 . (11)
. (11)
חשוב לציין כי המשוואות נתון חלים במקרה האידיאלי של גלאי ספירת פוטון אמיתי. עבור מערכות זיהוי אנלוגי, המשוואות הבאות חלות19,20:
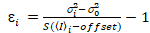 , (12).
, (12).
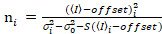 . (13)
. (13)
כאן, S הוא הגורם המרה בין פוטונים שזוהו הסעיפים דיגיטלי מוקלטות, 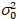 הוא רעש readout ואת היסט מתייחס ההיסט בעוצמה גלאי. באופן כללי, כמויות אלה צריכים להיות מכויל, עבור כל סוג הגלאי, המבוססת על מדידת השונות גלאי כפונקציה של עוצמת תאורה קבועה19, למשל, משטח מתכת רעיוני או פתרון דיי יבשים. היסט יכול להיקבע על ידי מדידת שיעור ספירה עבור דגימה ללא עירור אור. על ידי ביצוע רגרסיה ליניארית של השונות הקשורות גלאי
הוא רעש readout ואת היסט מתייחס ההיסט בעוצמה גלאי. באופן כללי, כמויות אלה צריכים להיות מכויל, עבור כל סוג הגלאי, המבוססת על מדידת השונות גלאי כפונקציה של עוצמת תאורה קבועה19, למשל, משטח מתכת רעיוני או פתרון דיי יבשים. היסט יכול להיקבע על ידי מדידת שיעור ספירה עבור דגימה ללא עירור אור. על ידי ביצוע רגרסיה ליניארית של השונות הקשורות גלאי 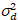 לעומת העוצמה (אני) העלילה, S ו יכול להיות נחוש19:
לעומת העוצמה (אני) העלילה, S ו יכול להיות נחוש19:
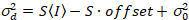 . (14)
. (14)
בסופו של דבר, הבהירות קרוס-המתאם מחושב בתוך כל פיקסל ולא מוגדר באופן כללי כמו21
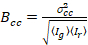 , (15).
, (15).
איפה 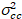 הוא קרוס-השונות
הוא קרוס-השונות 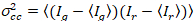 .
.
על מנת לסנן תנודות חיים ארוכים, כל ccN & B החישובים מבוצעים בעקבות קרון מטען סינון, באופן עצמאי עבור כל פיקסל22. בקצרה, ni, חדוהאני (אני = g, r) ו- Bcc מחושבים בהזזה מקטעים של למשל, 8-15 מסגרות. הערכים שהושג ובכך להיות בממוצע אז כדי להשיג הפיקסל האחרון מספר וערכי בהירות.
ניתוח סטויכיומטריה
כדי לאמוד את סטויכיומטריה של חלבון מתחמי תא-תא באנשי קשר, הבהירות מולקולרית ניתן בנפרד לנתח בכל אחד מהערוצים ספקטרליות עבור sFCCS או ccN & B נתונים. ב- sFCCS, מתקבל ערך הבהירות אחד לכל מדידה בכל ערוץ. ב ccN & B, היסטוגרמה הבהירות של פיקסלים כל הקשר תא-תא מתקבל, הערך הממוצע (או החציוני) יכול לשמש נציג בהירות למדידה. על ידי ביצוע הניתוח אותו על הפניה monomeric, כל ערכי הבהירות יכול להיות מנורמל להשיג ישירות מדינת oligomeric הממוצע מתחמי חלבון שזוהו. בנקודה זו, חשוב לתקן לנוכחות של FPs שאינו-פלורסנט שעלולים לגרום underestimation המדינה oligomeric. זה מתבצע בדרך כלל על ידי מודדים את הבהירות הפניה ההומו-dimeric חלבון23,24 באמצעות sFCS צבע אחד או מספר והבהירות (N & B).
