Isolation of stromal vascular fraction from human PVAT
Figure 1A shows a schematic of the anatomical region where the PVAT overlying the ascending aorta was obtained. We previously described the patient populations undergoing coronary artery bypass grafting from which these samples were derived6. Figure 1B shows an example of the human PVAT obtained following surgery. Figure 1C shows a representative PVAT tissue section stained with Masson’s Trichrome stain. Figure 1D is a phase contract micrograph showing a population of PVAT-derived stromal cells during the expansion phase prior to differentiation. Cells can be expanded and frozen for future use. Cells are typically frozen at a density of 2.5 x 105 cells/mL in a media comprised of 70% FBS, 20% basal (antibiotic free) DMEM F12 and 10% DMSO in an isopropanol cryochamber at -80 °C for 24 h and moved to the liquid phase of a liquid N2 freezer for long-term storage.
Adipogenic differentiation
Studies were performed in parallel with human bone marrow-derived MSC (Figure 2A,B) and PVAT-derived progenitor cells (Figure 2C,D). The left panels of Figure 2 show the non-induced condition, where no lipid accumulation is evident. The right panels show cells following adipocyte differentiation and staining of neutral lipids with Oil Red O. While the degree of differentiation in the human aortic PVAT-derived cells is more robust, both human cell sources exhibited the ability to differentiate towards the adipogenic lineage.
Osteogenic differentiation
The osteogenic differentiation protocol was used for human bone marrow-derived MSC (Figure 3A–C) and PVAT-derived cells (Figure 3D–F). Non-induced cells (Figure 3A,D) did not stain with Alizarin Red. After the osteogenic differentiation protocol, the human MSC developed calcified nodules that stained with Alizarin Red (Figure 3B–C), while human aortic PVAT-derived cells did not (Figure 3E–F). These data indicate that our preparation of cells from the stromal vascular fraction of human PVAT lack significant number of progenitors with the ability to undergo osteogenesis. Depending on the study and time course of differentiation, it is advisable to follow up standard stains with detection of molecular markers that define osteogenic lineage commitment (e.g. RUNX2, osterix, alkaline phosphatase) or osteoblasts (osteopontin, osteocalcin, alkaline phosphatase, BAP1).
Chondrogenic differentiation
Cells derived from both human bone marrow MSC (Figure 4A) and human PVAT (Figure 4B) display features characteristic of chondrogenic differentiation, with abundant collagen accumulation in the micromass. Micromasses formed from human bone marrow MSC and aortic PVAT-derived cells also exhibited abundant accumulation of glycosaminoglycans (blue) as indicated by Alcian blue staining (Figure 4C–D, respectively). Morphologically, structures similar to lacunae were detected with cells sitting in cavities surrounding by collagen deposition (Figure 4, arrows). Depending on the study and time course of differentiation, the detection of specific chondrogenic markers (aggrecan, collagen type II, osteonectin, Sox9) is useful.
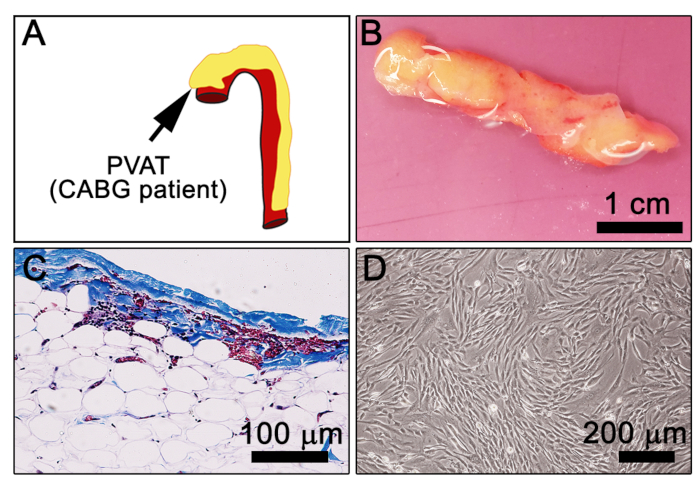
Figure 1: Morphological characteristics of human PVAT. (A) Cartoon depiction of the human aorta (red) with surrounding PVAT (yellow). Arrow indicates ascending aorta. (B) A 480 mg piece of human aortic PVAT from a CABG patient in DMEM prior to dissociation. (C) Masson’s trichrome staining of formalin-fixed paraffin-embedded human PVAT (dark brown/black = nuclei, blue/purple = connective tissue, pink = cytoplasm; Note, RBC appear reddish-pink. (D) Representative image of explanted PVAT-stromal cells at 7 days in culture. Please click here to view a larger version of this figure.
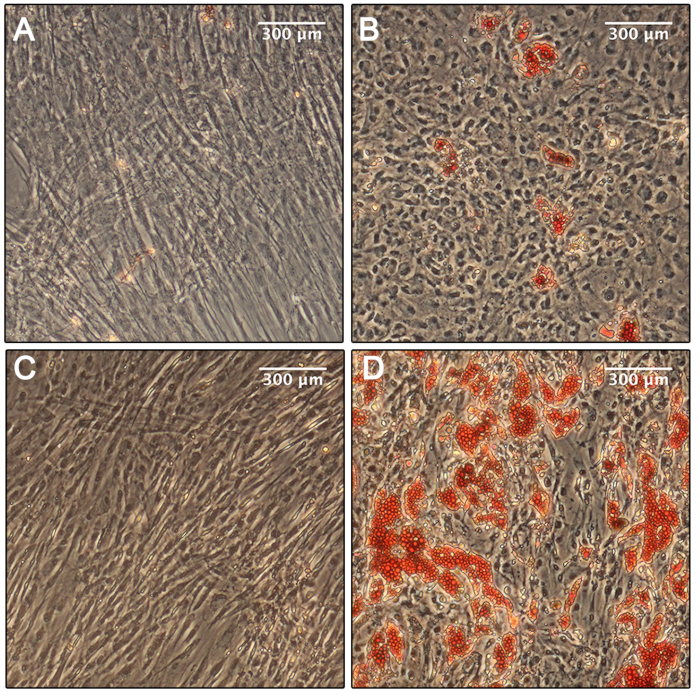
Figure 2: Adipogenic differentiation. (A) Phase microscopy of the human bone marrow MSC in the non-induced condition shows no evidence of differentiation. (B) Phase microscopy of the human bone marrow MSC in the induced adipogenic condition shows some differentiation and immobilization of neutral lipids, stained with Oil Red O after 14 days. (C) Phase microscopy of the human aortic PVAT-derived cells in the non-induced adipogenic condition shows no evidence of differentiation. (D) Phase microscopy of the human aortic PVAT-derived cells in the induced adipogenic condition shows robust differentiation and immobilization of neutral lipids, stained with Oil Red O after 14 days. Please click here to view a larger version of this figure.
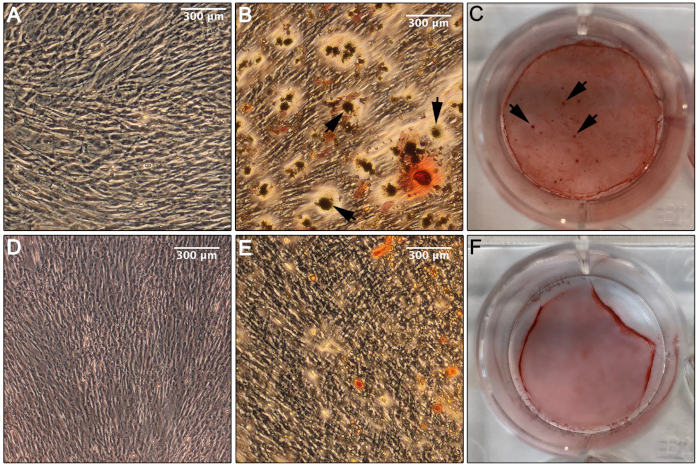
Figure 3: Osteogenic differentiation. (A) Phase microscopy of the human bone marrow MSC in the non-induced osteogenic condition shows no evidence of differentiation toward an osteogenic lineage. (B) Phase microscopy of the human bone marrow MSC in the induced condition after 14 days in culture shows evidence of differentiation and formation of calcium deposits stained with Alizarin Red (arrows). (C) An image of the well containing the human bone marrow MSC in the induced osteogenic condition following staining with Alizarin Red indicates abundant calcium deposition, suggesting successful differentiation toward an osteogenic lineage (arrows, calcium deposition). (D) Phase microscopy of the aortic human PVAT-derived cells in the non-induced osteogenic condition exhibits no indication of differentiation toward an osteogenic lineage. (E) Phase microscopy of the aortic human PVAT-derived cells in the induced osteogenic condition after 14 days in culture shows no evidence of differentiation toward an osteogenic lineage or deposition of calcium when stained with Alizarin Red, only non-specific staining. (F) An image of the well containing the human aortic PVAT-derived cells in the induced osteogenic condition following staining with Alizarin Red shows only non-specific staining. Please click here to view a larger version of this figure.
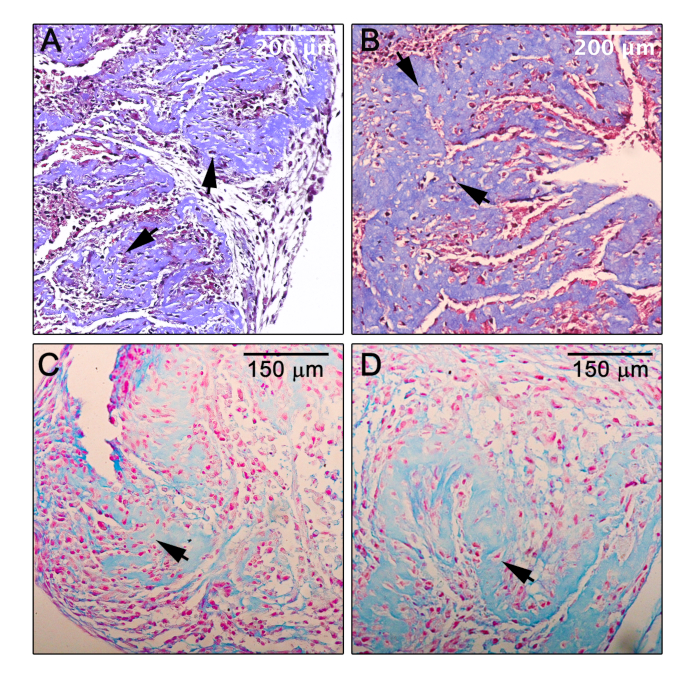
Figure 4: Chondrogenic differentiation. (A,B) Light microscopy of a section of the micromass formed by human bone marrow MSC (A) or human aortic PVAT progenitor cells (B) in the induced chondrogenic condition after 14 days in culture and subsequently stained with Masson’s Trichrome, which indicates significant deposition of collagen (blue) and suggests successful differentiation toward a chondrogenic lineage. (C,D) Light microscopy of a section of the micromass formed by human bone marrow MSC (C) or human aortic PVAT progenitor cells (D) in the induced chondrogenic condition after 14 days in culture and subsequently stained with Alcian blue, which indicates deposition of acidic proteoglycans (e.g. glycosaminoglycans) as are typically found in cartilage. Counterstain, nuclear fast red; arrows, lacunae-like structures. Please click here to view a larger version of this figure.
