הופעל מקושרת צולבת מקושרת לפיתוח מהיר של שרפים כרומטוגרפיה של אהדה-לכידת נוגדן כמחקר מקרה
Summary
בהליך זה, מבוסס על מעטפת המבוססת על DsRed-בסיס להפקת שרף מיוחד בררני ללכידת נוגדנים חד-שבטיים מתמציות צמחים גולמי או סופרנטנמלים של תרבות התאים, כחלופה לחלבון A.
Abstract
טיהור של נוגדנים חד שבטיים (mAbs) מושגת בדרך כלל על ידי חלבון כרומטוגרפיה זיקה, אשר יכול להסביר עד 25% מעלויות התהליך הכולל. אלטרנטיבי, שלבי הלכידה החסכונית הם אפוא רב ערך עבור ייצור בקנה מידה תעשייתי, שבו כמויות גדולות של mAb יחיד מיוצרים. כאן אנו מציגים שיטה עבור השתק של האפיפי המבוססת על הצלב האדום המבוסס על שרף משני מקושר המאפשר לכידת סלקטיבית של הנוגדן לנטרול האיידס 2F5 מפני תמציות צמחים גולמי מבלי להשתמש בחלבון A. האפיפי הליניארי ELDKWA היה התמזגו לראשונה גנטית לחלבון הפלורסנט DsRed וחלבון ההיתוך התבטא בטבק טרנסגניים (טבק טבאקו) צמחים לפני טיהור על ידי קיבוע מתכת-יון כרומטוגרפיה זיקה. יתר על כן, שיטה המבוססת על התקפה צולבת מקושרת שהופעלה הייתה ממוטבת לדחיסות גבוהה, לצימוד יעיל ולעלויות נמוכות. הרכב ה-pH והאגירה והריכוז המיסיסים היו הפרמטרים החשובים ביותר במהלך הליך הצימוד, ששופר באמצעות גישת עיצוב-ניסויים. שרף האהדה שנוצר נבדק על יכולתה לאגד באופן סלקטיבי את mAb היעד בתמצית צמח גולמי המאגר ממוטב עבור התאוששות גבוהה mAb, פעילות המוצר ויציבות שרף אהדה. השיטה יכולה בקלות להיות מותאמת לנוגדנים אחרים עם אפיסקופים ליניאריים. האפוקסידיות החדשות מאפשרות הגבלות על המצב העדין יותר מאשר חלבון A ויכול גם להקטין את העלויות של שלב הלכידה הראשונית עבור ייצור mAb.
Introduction
המוצרים הביופרמצבטיקה חשובים לטיפול בספקטרום רחב של מחלות כמעט בכל ענף של רפואה1. נוגדנים חד שבטיים (mAbs) להשתלט על השוק הביופרמקולוגיה, עם מכירות ברחבי העולם צפויים להגיע כמעט €110,000,000,000 בתוך 20202. פלטפורמת הביטוי המועדף על mabs הם הסינית שחלות אוגר תאים קווי, אשר בדרך כלל לייצר גבוה mabs מגדל של עד 10 g ∙ L-1 בתרבות supernatant3,4. עם זאת, הייצור של mAbs בתרביות תאים היונקים הוא יקר בשל העלות הגבוהה של המדיום ואת הצורך תסיסה סטרילית5. פלטפורמות ביטוי חלופיות כגון צמחים העשויים להציע גישה מהירה יותר, פשוטה יותר, זולה יותר ומדרגית יותר עבור ייצור תעשייתי6,7.
בנוסף לעלויות הקשורות לתרבויות תאי היונקים, השימוש הנרחב בחלבון כרומטוגרפיה של זיקה ללכידת המוצר הוא מנהל התקן של עלות מרכזית עבור הייצור התעשייתי של mAbs. חלבון A נמצא באופן טבעי על פני השטח של התאים האורהוניים של סטיילולוקוקוס והוא מאגד את הרסיס crystallizable (Fc) באזור של murine מסוימים ונוגדנים אנושיים, ובכך מתנהג כמנגנון הגנה להתחמק המערכת החיסונית ומוראלית8. חלבון A הפך לתקן זהב תעשיית לכידת mAbs מן התרבות התאית סופר והוא גם בשימוש נרחב על ידי קהילת המחקר כי זה סלקטיבי מאוד, בדרך כלל להשיג Mabs purities של ~ 95% בשלב אחד8. באופן לא מפתיע, מכירות של חלבון A במהלך שני העשורים האחרונים יש שיקוף מקרוב את המכירות של mAbs8. בהתאם לסולם הייצור, העלויות של חלבון A יכול להתאים יותר 25% מעלויות התהליך הכולל ובכך להשפיע על מחיר השוק של mabs טיפולית, אשר יכול להיות עד €2,000 g-15,9. לכן, כרומטוגרפיה חלופית שרפים עם ביצועי טיהור דומה יש פוטנציאל להפחית באופן משמעותי את עלויות הייצור, ביצוע טיפולים נוגדנים מבוססי נגיש עבור מספר גדול יותר של חולים10,11 ,12. חלופות כאלה עלולות גם לעקוף את החסרונות של חלבון כרומטוגרפיה, כולל התנאים הקשים ביותר ב-pH נמוך (בדרך כלל < 3.5) העלולים לגרום ל-mAbs לעבור שינויים מתוניים המקדמים את הצבירה13 . וחשוב מכך, חלבון A סלקטיבי רק עבור אזור Fc של מחלקות IgG מסוימות, כך שאינם פונקציונליים מולקולות עם תחומים כריכה חתוכים עשוי לטהר עם המוצר ללא שינוי5, ואילו mab ליגוניות כגון קטעי שרשרת בודדת משתנה לא להיקשר לחלבון A בכלל.
כאן, אנו מתארים שרף כרומטוגרפיה של זיקה כרומטוגרפית ללכידת ה-HIV-נטרול mab 2f5 באמצעות האפיפפן הליניארי שלה eldkwa (אות אחת חומצת אמינו קוד)5,14. התחזרנו גנטית את האפיפי של 2F5 ל-C הטרמינוס של חלבון הפלורסנט DsRed, אשר פעלו כמולקולה המוביל והעיתונאי, והפיק את החלבון המתקבל Dsred-2F5-Epitope (dfe) בטבק הטרנסגניים ( צמחים טבק טבאקו). DFE היה מטוהר על ידי צעד אחד מקיבוע מתכת-יון זיקה כרומטוגרפיה (IMAC). השתק של האהדה הטהורה של הזיקה ליגולי אל שרף אגקם מקושר הושג על ידי צימוד כימי באמצעות N-הידרוקסיזינימיד (הארגון)-הופעלו מקושרות מוצלבות עמודים. עיצובים ניסיוניים סטטיסטיים שימשו אז כדי למטב את הליך השתק ויעילות צימוד15. אסטרטגיית טיהור עבור mAb 2F5 הוערך במונחים של טוהר נוגדנים, תשואה ליגויציבות. בניגוד לחלבון A, אשר מאגד את אזור Fc, DFE מאוגד לאזור הקבוע ביותר של 2F5, להבטיח טיהור של מולקולות עם paratope שלם. הקונספט שלנו יכול בקלות להיות מותאם לכל mAb עם אפירופה ליניארי או אחרים מבוססי פפטיד מבוסס ליגנדס אשר ניתן לזהות בקלות על ידי מחקרים microarray16.
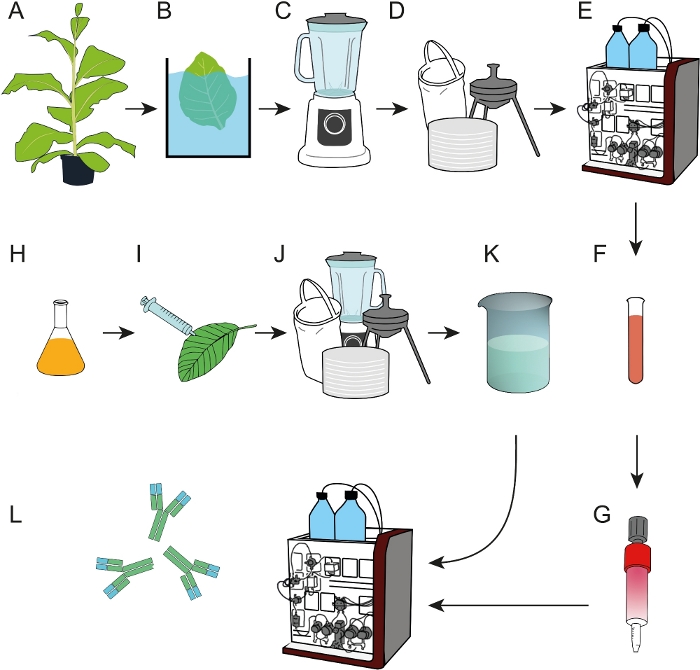
איור 1: תהליך תזרים התהליכים להכנת שרפים אהדה אפירופה שניתן להשתמש בהם ללכידת mAbs מתמציות צמחים גולמי או סופרנטנמלים של תרבות התאים. (א) ליגו dfe האהדה התבטא במפעלי טבק טרנסגניים. צעד משקעים בחום (ב) נכלל לפני העלים שנקטפו היו הומוגניים (C). (ד) תמצית הצמח הגולמי הובהר על ידי סינון שקית, סינון עומק ו-0.2 יקרומטר סינון סטרילי. (ה) dfe טוהר לאחר מכן על ידי IMAC. (F, G) האהדה DFE מטוהרים ליגס היה מחובר על העמודים EDC/הפעילה החוצה המופעלות מחוצה. (ח) בתרבויות חיידקית הנושאות את הנוגדן לקידוד T-DNA 2F5 שימשו לביטוי חולף ב -N. benthamiana צמחים (I) גדל בפיטוטרון. (J) N. העלים נקצרו ועובדו כמתואר ב D. (K) mab 2f5 טוהר מן התמצית הבהיר באמצעות עמודות זיקה dfe (L). אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
Protocol
Representative Results
Discussion
יישומים של שרף הזיקה הרומן
הצגנו כי מותאם אישית הזיקה כרומטוגרפיה שרפים ללכידת mAbs יכול להיות מיוצר על ידי השתק ליגייט והמכיל אפירופה ספציפי Mabs לתוך הפעיל מקושרת מקושרת. כדי לעצב שרף כזה, היה צורך לדעת את רצף האפירופה ולהשתמש באפירופה לינארית. שרפים וכתוצאה מכך הם יתרון עבור לכידת mAbs כי הם יכולים להחליף חלבון יקר שלבים כרומטוגרפיה זיקה. האינטראקציה בין 2F5 ו-DFE במחקר שלנו היה בתיווך על ידי אפירופה – paratope כריכה, כך ליגאן שלנו צריך להיות סלקטיבי יותר מאשר חלבון A, אשר נקשר לאזור Fc של רוב מורלין והאיגים אנושיים. מכיוון שליגניות בודדות נדרשות עבור כל mAb, השיטה שלנו עשויה להיראות בתחילה מתאימה בעיקר לנוגדנים המיוצרים בקנה מידה גדול מאוד. עם זאת, על ידי שילוב הגישה שלנו עם ביטוי מהיר המבוסס על הצמח בחלבון ארעי, ליגנדס אהדה חדשה יכול להיות מוכן בתוך פחות מ 2 שבועות27 עם מאמץ מינימלי28. מכאן, השיטה מתאימה גם לטיהור בקנה מידה קטן mAb.
הפקה ושיפורים פוטנציאליים של הזיקה ליגולי
הצמחים מציעים פלטפורמת ייצור מהירה ובטוחה עבור ליגטרים5,29,30, כגון חלבון היתוך dfe מובלט במחקר שלנו מקרה. חולצ’ינג חומר הצמח הפחית במידה ניכרת את כמות החלבונים של תא מארח בצעד אחד והיה משתלב בקלות בשגרת הבהרה סטנדרטית. עם זאת, ההתאוששות של ליגנד היה נמוך ההתקנה הנוכחית, כנראה בשל היציבות תרמית מתון שלה כריכה מסוימת לא ספציפית לשכבות מסנן, כפי שדווחו עבור מוצרים אחרים31,32,33. הנדסה המוביל כדי להגדיל את היציבות התרמית שלה ולכן לסייע לשפר את התפוקה בעתיד, כפי שמתואר עבור המועמד החיסון נגד מלריה CCT, אנזים antitumor של הגידול או מβ lic-msm-glucosidase34, 35,36. אותו הדבר נכון גם לגבי שלב סינון העומק, שבו הנדסת חלבונים עשויים לסייע להפחית את הכריכה הלא ספציפית לחומר המסנן37. עלויות הייצור עבור dfe וליגטים דומים יכולים גם להיות מופחת על ידי שיפור היעילות הכוללת של הבהרה באמצעות flocculants או מסנן תוספים38,39.
כאשר DsRed משמש כמוביל, הוא יוצר קומפלקס tetrameric. זה יתרון כי זה מגדיל את מספר האפיסקופים ללימס, אבל זה עשוי גם להפוך את הליגולי רגישים יותר לפירוק או הדנטורציה במהלך כרומטוגרפיה של זיקה. חלבון הנושאת monomeric כגון mcherry יכול להיות עדיף, כי זה יציב ב-pH נמוך40, והכללת חוזר דו-מושביים יהיה להגדיל את האוויקות של ליגנד ובכך להגדיל את קיבולת שרף5, מיכל בן 26 , 41. מערכות הייצור הפשוטות של המוביל (כלומר, לאלה שאין להם קשרים דיאטוניים או שינויים שלאחר ההעברה) עשויים להפחית את עלויות הייצור ולהפוך את הליגשות לתחרותיות יותר בחלבון A. לדוגמה, חלבון פלורסנט ירוק כבר ביטא בתאים חיידקיים עם תשואה של ~ 1 גרם · ק”ג-1 ביומסה, אשר להפחית באופן משמעותי ליגוהייצור עלויות42.
ללא קשר למארח הביטוי, משיכה מטוהרים ליגורה היה נדרש במהלך הצימוד כדי למזער את השתק של חלבונים תא מארח או רכיבי מדיה שיכולים אחרת להפחית את הסלקטיביות שרף וקיבולת. הכללת תג פולי histidine עבור טיהור IMAC הגביר את הטוהר כדי ~ 90% בשלב אחד, הקלה ליגנד מהירה וזולה ייצור5,43,44. עם זאת, מיקומו של תג ההיתוך חשוב משום שיש לו את הפוטנציאל לעכב את הכריכה הקוסמית או לגרום למחשוף של התג או האפירופה מהמוביל45,46.
שינזציה של הזיקה לליבטורים בעלי כרומטוגרפיה מופעלים
השתק בוצע באופן ידני או באמצעות מערכת כרומטוגרפיה. אמצעי האחסון של המאגר הקטן לעמודה נראו כבעד טיפול ידני (לדוגמה, עקב אמצעי האחסון המינימלי לפסולת). עם זאת, אם יש צורך בעמודות מרובות/יותר, מערכת הכרומטוגרפיה הופכת את תנאי הצימוד לקל יותר לשליטה (לדוגמה, שיעורי זרימה מוסדרים) ולכן סביר יותר להשיג תוצאות שונות במונחים של DBC. הנתונים שלנו מראים כי מאגר צימוד ו-pH יש השפעה חשובה על היעילות צימוד ועלויות העמודה הכוללת. גורמי הקרנה המשפיעים על תגובת הצימוד והתאמת החיבור שלהם לכל חלבון מוביל (או אפילו עבור כל נשאית-ליגיון) יכולים אפוא לשפר את יעילות הצימוד והביצועים של שרף, ואנו ממליצים על גישה זו.
בדיקת בידוד 2F5 באמצעות שרף של זיקה DFE
תשואה המוצר וטוהר הם היבטים חשובים של ביצועים שרף, ובמקרה של DFE השגנו תשואה של 105 ± 11% (± SD, n = 3) וטוהר של 97 ± 3% (± SD, n = 3), אשר דומה לביצועים של חלבון בחינת ביצועים שרפים25 ,26. עוד מחוון ביצועים מפתח עבור שרפים בכלל (ובמיוחד עבור אלה המבוססים על ליגניות אהדה) הוא DBC ב 10% פריצת מוצר, כי פרמטר זה משפיע על כמות השרף הנדרש עבור תהליך ספציפי ולכן את העלויות. עבור הליגאות DFE, DFE הראשונית היה ~ 4 g · L-1 שרף, שהוא ~ 13% מהערך המתאים עבור חלבון A בתנאים דומים (רק 2 זמן יצירת קשר)25,47 אבל על 15-קיפול גבוה יותר לעומת שרפים זיקה מותאם אישית אחרים כגון אנטי-FSH-זיקה חיסוני ליגו שימוש באותו זמן מגורים של 2 דקות48. DBC של DBC סירב 15% מהערך ההתחלתי לאחר 25 מחזורי bind ו-elute ואילו יותר מ 50 מחזורים נדרשים לאותו אובדן של DBC בחלבון מסחרי שרפים49. עם זאת, חשוב לציין כי המוביל שלנו עדיין לא היה מותאם באותה מידה כמו חלבון A, אשר נחקר באופן מקיף והשתפר בארבעת העשורים האחרונים8.
עד כה שיפרנו את היציבות שרף והתאוששות המוצר על ידי מעבר מ-pH נמוך מאגר לריכוז גבוה של מגנזיום כלוריד (איור 3), כמומלץ במחקרים קודמים13. הצבע האדום האופייני של ליגולי האהדה לא נמוג באופן משמעותי במהלך 25 לאגד-ו-elute מחזורים, כך אנו משערים כי הצמח האנדוסוגני בתמציות צמחים הבהיר31 אולי נחתך ובכך להשבית את האפירופה של הליו. לכן, עיצוב מחברים עמידים פרוטאז כדי לחבר את המנשא ואת האפירופה עשוי לסייע לשמור על DBC הראשונית על מספר מורחב של מחזורים. בהינתן הביטוי המהיר והפשוט והטיהור של הליגאפה DFE, הזיווג הישיר שלה לגבי שרפים כרומטוגרפיה מסחריים, והתשואה המעולה שלה למוצר וטוהר, אנו מאמינים כי השיטה שלנו מציעה חלופה מתאימה חלבון A עבור טיהור של mAbs ונגזרות נוגדן אשר לא לאגד חלבון A, במיוחד אם שיפורים המוביל והמקשר יכול לשפר את DBC ו ליגויציבות. הנחה זו היתה נתמכת על ידי ההבדל הקטן בקבוע הדיסוציאציה של DFE מטוהרים וחלבון A-מטוהרים 2F5 נוגדן5, המציין כי ליגתנו החדש זיקה מאפשר התאוששות mabs באיכות גבוהה.
הטבות ומגבלות נוכחיות של השיטה
הפקת ליגולי אהדה כמו היתוך גנטי עם הספק חלבון מגביר מסיסות במאגרים מימית ובכך תאימות עם התנאים האופייניים לליגנד צימוד. לעומת זאת, פפטידים ריקים נגזר סינתזה פפטיד הפאזה היציבה עשויה להיות מוגבלת מסיסות תחת תנאים אלה בשל הרצף שלהם50, אשר לא ניתן לשנות כי זה מוכתב על ידי רצף חומצת אמינו של אפירופה מוכר על ידי mab כדי ל לטוהר. אחרים השתמשו לפיכך סינתזה על שרף של פפטיד ליגנדס51. יכולת האיגוד הסטטי של שרף שנוצר היה גבוה (~ 80 g · L-1), אך תהליך ההכנה של שרף הוא ארוך, קיבולת מחייבת דינמית לא דווחה והטוהר וההחלמה שהושגו היו נמוכים מאשר בגישה שלנו. יתרון נוסף של חלבון היתוך ליגולי בקנה מידה הוא כי הליקלים והמשתנים שלהם יכול להיות מיוצר במהירות, מטוהרים ונבדק במאמץ מינימלי קל לשימוש מערכת ביטויים גבוהה-דרך52.
שתי המגבלות הנוכחיות של השיטה המוצגת כאן הן קיבולת הכריכה הדינמית הנמוכה של 3 גר’ · L-1 והפחתת 90% שלה במהלך 25 מחזורי bind ו-elute5. ניתן לטפל במגבלות אלה בעתיד על-ידי החלת תנאי טעינה מחמירים פחות והחלפת הספק הנוכחי של DsRed באמצעות variant מהונדס ויציב יותר בהתאמה. לדוגמה, הכפלה של זמן יצירת הקשר הנוכחי מ-2 עד 4 דקות כוללת את הפוטנציאל להכפיל את קיבולת האיגוד הדינאמי כפי שהוצג עבור חלבון מסוים שרפים26.
פתרון בעיות
הטבלה הבאה מדגישה בעיות פוטנציאליות שבהן ניתן להיתקל במהלך פרוטוקול זה ומספקת רמזים על אופן הפתרונן (טבלה 1).
| טבלה 1: בעיות פוטנציאליות שניתן להיתקל בהן ותיקונים אפשריים. | |||
| שלב הפרוטוקול | עיה | וודאי | תקן |
| 1 | הצמחים לא גדלים | תנאי הגדילה הפרוצים | בדוק את ה-pH ואת המוליכות של הדשן |
| בדוק את הטמפרטורה ואת תנאי האור | |||
| 2 ו-3 | כמויות גדולות של חלבונים תא מארח נמצאים לאחר החילוץ | משקעים לא גמורים | בדוק את הטמפרטורה במהלך השמיכה |
| בדוק את העצבנות באמבט השמיכה | |||
| 2 ו-3 | לא נמצא מוצר בתמצית הצמח | הטמפרטורה החולצ’ינג גבוהה מדי | בדיקת טמפרטורה pH במהלך השמיכה |
| pH במאגר החולטים נמוך מדי | |||
| 3 | חלקי גבעול או עלים גדולים נותרים לאחר החילוץ | ערבוב לא שלם בבלנדר | ודא שחומר הצמח אינו יוצר תקע בבלנדר |
| 3 | עלייה בלחץ מהיר במהלך סינון עומק | בחירת מסננים שגויה ו/או כיוון | בדיקת סוג המסנן וכיוון הסינון |
| 4 | חלבון היתוך קטן במהלך הימנעות/הרבה חלבון היתוך בזרימה-דרך | שרף IMAC לא הואשם ביוני מתכת | בדוק אם שרף ה-IMAC הואשם כהלכה ביונים |
| חלבון היתוך איבד את התגית הקרבה | הימנע מאור השמש האינטנסיבי וטמפרטורות גבוהות במהלך טיפוח הצמח | ||
| 4 | חלבון פיוז’ן שאבד במהלך הריכוז | חלבון פיוז’ן מאוגד לקרום | בדוק את סוג הממברנה |
| ודא שגורם הריכוז לא היה גבוה מדי | |||
| מיכל 5 | תשואה נמוכה לצימוד | רצף שגוי של תוספת מגיב צימוד | בדוק את התוויות הריאגנטים ואת רצף החיבור |
| הכנה שגויה של העמודות לפני צימוד | בדוק את התנאים של הוראות העמודה | ||
| 5 ו-6 | תשואה נמוכה של mAb | ביטוי נמוך מ-mAb ביומסה הצמח | מבחן mAb ביטוי ביומסה |
| ליגוצפיפות נמוכה | בדוק את טוהר הכנת חלבון ההיתוך | ||
| 7 | ריכוז נמוך/חלבון גבוה מאוד בתוך שיטת ברדפורד | היווצרות בועות במהלך העוגה | בדוק אם בועות 96-ובכן palte |
| 7 | ריכוז נמוך של mAb במהלך SPR מדידה | חלבון שנפרץ שבב | השווה עם תוצאות של mAb סטנדרטי עם ריכוז ידוע |
| דילול לדוגמה שגויה | בדיקת שיעור הדילול והחוצץ | ||
שולחן 1: בעיות הירי.
Disclosures
The authors have nothing to disclose.
Acknowledgements
היינו רוצים להכיר באיברהים אל-עמאדי על הטיפוח של צמחי הטבק הטרנסגניים וד ר תומס רדמאכר על מתן ביטוי הטבק וקטור. המחברים רוצים להודות לד ר ריצ’רד טוימאן לקבלת סיוע מערכתי ומרקוס סאק לדיונים פוריים על מבנה הזיקה של DFE. העבודה הזאת ממומנת בחלקו על ידי התוכניות הפנימיות של פראונהופר-גיסלסירכתיים תחת גרנט לא. מושכים 125-600164 ומדינת צפון-ריין-וסטפאליה תחת מענק לייאסטגשיום מעניק no. 423 “רשת, הפקה מסתגלת”. עבודה זו נתמכה על ידי הגרמני Forsch, החברה במסגרת מערכת הכשרת מחקר “מסירת תרופות ממוקדות גידול” גרנט 331065168. ביטוח הבריאות של GE תמך בפרסום הגישה הפתוחה של מאמר זה.
Materials
| 10L/20L Bucket | n/a | n/a | Blanching equipment |
| 2100P Portable Turbidimeter | Hach | 4650000 | Turbidimeter |
| ÄKTApure | GE Helthcare | 29018226 | Chromatography system |
| Allegra 25R | Beckman Coulter | 369434 | Centrifuge |
| Amine Coupling Kit | GE Healthcare | BR100050 | SPR chip coupling kit |
| Amine Coupling Kit | GE Healthcare | BR100050 | SPR chip coupling kit |
| Antibody 2G12 | Fraunhofer IME | n/a | Standard for SPR quantification |
| Blender | Waring | 800EG | Blender |
| BP-410 | Fuhr | 2632410001 | Bag filter |
| CanoScan 5600F | Canon | 2925B009 | Scanner |
| Centrifuge tube 50 mL self-standing | Labomedic | 1110504 | Reaction tube |
| Chelating Sepharose FF | GE Helthcare | 17-0575-01 | Chromatography resin |
| Cond 3320 | WTW | EKA 3338 | Conductometer |
| Design-Expert(R) 8 | Stat-Ease, Inc. | n/a | DoE software |
| Discovery Compfort | Gilson | F81029 | Multichannel pipette |
| Disodium phosphate | Carl Roth GmbH | 4984.3 | Media component |
| Diverse bottles | Schott Duran | n/a | Glas bottles |
| Dri Block DB8 | Techne | Z381373 | Heat block |
| DsRed | Fraunhofe IME | n/a | Standart |
| EDTA | Carl Roth GmbH | 8043.2 | Buffer component |
| EnSpire | Perkin Elmer | 2390-0000 | Plate reader |
| ETHG-912 | Oregon Scientific | 086L001499-230 | Thermometer |
| F9-C | GE Helthcare | 29027743 | Fraction collector |
| Ferty 2 Mega | Kammlott | 5.220072 | Fertilizer |
| Forma -86C ULT freezer | ThermoFisher | 88400 | Freezer |
| HEPES | Carl Roth GmbH | 9105.3 | Buffer component |
| Hettich Centrifuge Mikro 200 | Hettich | 2400 | Centrifuge |
| HiPrep 26/10 | GE Helthcare | GE17-5087-01 | Chromtography column |
| HiTrap NHS-activated Sepharose HP, 1 mL | GE Helthcare | 17-0716-01 | Chromatography columns |
| Hydrochloric acid | Carl Roth GmbH | 4625.1 | Buffer component |
| Imidazole | Carl Roth GmbH | 3899.2 | Buffer component |
| K700 | Pall | 5302305 | Depth filter layer |
| KM02 basic | IKA | n/a | Magnetic stirrer |
| KS50P 60D | Pall | B12486 | Depth filter layer |
| L/S 24 | Masterflex | SN-06508-24 | Tubing |
| Lauda E300 | Lauda Dr Wobser GmbH | Z90010 | Immersion circulator |
| Magnesium chloride | Carl Roth GmbH | KK36.2 | Buffer component |
| Masterflex L/S | Masterflex | HV-77921-75 | Peristaltic pump |
| Minisart 0.2 µm | Sartorius | 16534K | Filter unit |
| Nalgene Rapid-Flow PES bottle top filter | Thermo Fischer Scientific | 595-4520 | Vacuum filtration of SPR buffers |
| Nickel sulphate | Carl Roth GmbH | T111.1 | Buffer component |
| Novex NuPAGE 4-12% BisTris LDS gels | Invitrogen | NP0336BOX | LDS-PAA gels |
| Novex X-cell Mini Cell | Invitrogen | EI0001 | PAGE chamber |
| NuPAGE 20x running buffer | Invitrogen | NP0002 | Buffer concentrate |
| NuPAGE antioxidant | Invitrogen | NP0005 | Antioxidant |
| PageRuler protein ladder (10-180 kDa) | Invitrogen | 26616 | Protein standart |
| Perforated bucked | n/a | n/a | Blanching |
| PH 3110 | WTW | 2AA110 | PH meter |
| PowerPac HC | Biorad | 1645052 | Electrophoresis module |
| Protein A from Staphylococcus aureus | Sigma-Aldrich | P7837-5MG | Coating of SPR chips |
| Sephadex G-25 fine, cross linked dextran | GE Helthcare | 17003301 | Chromatography resin |
| Silicone spoon | n/a | n/a | Spoon |
| Simply Blue SafeStain | Invitrogen | LC6060 | Gel staining solution |
| Sodium acetate | Carl Roth GmbH | 6773.1 | Buffer component |
| Sodium acetate | Carl Roth GmbH | X891.1 | Media component |
| Sodium azide | Sigma Aldrich | S2002-100G | Media component |
| Sodium chloride | Carl Roth GmbH | P029.2 | Buffer component |
| Sodium citrate | Carl Roth GmbH | HN13.2 | Buffer component |
| Sodium bisulfite | Carl Roth GmbH | 243973-100G | Media component |
| Sodium phosophate | Carl Roth GmbH | T877.2 | Media component |
| SPR Affinity Sensor – High Capacity Amine | Sierra Sensors GmbH/Bruker Daltonics | SPR-AS-HCA | SPR chip |
| SPR-2/4 Surface Plasmon Resonance Analyzer | Sierra Sensors GmbH/Bruker Daltonics | n/a | SPR device |
| SSM3 | Stuart | 10034264 | Mini Gyro-rocker |
| Heated vessel, 20 L | Clatronic | n/a | Blanching chamber |
| Sterile syringes, 2 mL | B. Braun | 4606027V | Syringes |
| Syringe adpter (Union Luer F) | GE Helthcare | 181112-51 | Syringe adapter |
| TE6101 | Sartorius | TE6101 | Precision scale |
| Tween-20 (Polysorbate) | Merck | 8170721000 | Buffer component |
| Unicorn 6.4 | GE Helthcare | 29056102 | Chromatography software |
| Vacuum bags | Ikea | 203.392.84 | Plant storge |
| VelaPad 60 | Pall | VP60G03KNH4 | Filter housing |
| Vortex-Genie 2 | Scientific industries | SI-0236 | Vortex |
| XK-26/20 column housing | GE Helthcare | 28-9889-48 | Chromtography column |
References
- Kesik-Brodacka, M. Progress in biopharmaceutical development. Biotechnology and Applied Biochemistry. 65 (3), 306-322 (2018).
- Ecker, D. M., Jones, S. D., Levine, H. L. The therapeutic monoclonal antibody market. MAbs. 7 (1), 9-14 (2015).
- Jayapal, K., Wlaschin, K. F., Hu, W. S., Yap, M. G. S. Recombinant Protein Therapeutics from CHO Cells – 20 Years and Counting. Chemical Engineering Progress. 103, 40-47 (2007).
- Kunert, R., Reinhart, D. Advances in recombinant antibody manufacturing. Applied Microbiology and Biotechnology. 100 (8), 3451-3461 (2016).
- Rühl, C., Knödler, M., Opdensteinen, P., Buyel, J. F. A linear epitope coupled to DsRed provides an affinity ligand for the capture of monoclonal antibodies. Journal of Chromatography A. 1571, 55-64 (2018).
- Edgue, G., Twyman, R. M., Beiss, V., Fischer, R., Sack, M. Antibodies from plants for bionanomaterials. Nanomedicine and Nanobiotechnology. 9 (6), e1462 (2017).
- Buyel, J. F., Fischer, R. Very-large-scale production of antibodies in plants: The biologization of manufacturing. Biotechnology Advances. 35 (4), 458-465 (2017).
- Bolton, G. R., Mehta, K. K. The role of more than 40 years of improvement in protein A chromatography in the growth of the therapeutic antibody industry. Biotechnology Progress. 32 (5), 1193-1202 (2016).
- Kelley, B. Industrialization of mAb production technology: the bioprocessing industry at a crossroads. MAbs. 1 (5), 443-452 (2009).
- Brochier, V., Ravault, V. High throughput development of a non protein A monoclonal antibody purification process using mini-columns and bio-layer interferometry. Engineering in Life Sciences. 16 (2), 152-159 (2016).
- Arakawa, T., Futatsumori-Sugai, M., Tsumoto, K., Kita, Y., Sato, H., Ejima, D. MEP HyperCel chromatography II: binding, washing and elution. Protein Expression and Purification. 71 (2), 168-173 (2010).
- Barroso, T. B., Aguiar-Ricardo, R. J., Roque, A. C. Structural evaluation of an alternative Protein A biomimetic ligand for antibody purification. Journal of Computer-aided Molecular Design. 28 (1), 25-34 (2014).
- Mazzer, A. R., Perraud, X., Halley, J., O’Hara, J., Bracewell, D. G. Protein A chromatography increases monoclonal antibody aggregation rate during subsequent low pH virus inactivation hold. Journal of Chromatography A. 1415, 83-90 (2015).
- Sack, M., et al. Functional analysis of the broadly neutralizing human anti-HIV-1 antibody 2F5 produced in transgenic BY-2 suspension cultures. FASEB Journal. 21 (8), 1655-1664 (2007).
- Buyel, J. F., Fischer, R. Characterization of Complex Systems Using the Design of Experiments Approach: Transient Protein Expression in Tobacco as a Case Study. Journal of Visualized Experiments. (83), 51216 (2014).
- Trasatti, J. P., Woo, J., Ladiwala, A., Cramer, S., Karande, P. Rational design of peptide affinity ligands for the purification of therapeutic enzymes. Biotechnology Progress. 34 (4), 987-998 (2018).
- Buyel, J. F., Gruchow, H. M., Boes, A., Fischer, R. Rational design of a host cell protein heat precipitation step simplifies the subsequent purification of recombinant proteins from tobacco. Biochemical Engineering Journal. 88, 162-170 (2014).
- Simonian, M. H., Smith, J. A. Spectrophotometric and colorimetric determination of protein concentration. Current Protocols in Molecular Biology. 76 (Chapter 10), (2006).
- Buyel, F. J., Kaever, T., Buyel, J., Fischer, R. Predictive models for the accumulation of a fluorescent marker protein in tobacco leaves according to the promoter/5’UTR combination. Biotchnology and Bioengeneering. 110 (2), 471-483 (2013).
- Piliarik, M., Vaisocherova, H., Homola, J. Surface plasmon resonance biosensing. Methods in Molecular Biology. 503, 65-88 (2009).
- Buyel, J. F., Fischer, R. Scale-down models to optimize a filter train for the downstream purification of recombinant pharmaceutical proteins produced in tobacco leaves. Biotechnology Journal. 9 (3), 415-425 (2014).
- Baird, G. S., Zacharias, D. A., Tsien, R. Y. Biochemistry, mutagenesis, and oligomerization of DsRed, a red fluorescent protein from coral. Proceedings of the National Academy of Sciences, USA. 97 (22), 11984-11989 (2000).
- Vrzheshch, P. V., Akovbian, N. A., Varfolomeyev, S. D., Verkhusha, V. V. Denaturation and partial renaturation of a tightly tetramerized DsRed protein under mildly acidic conditions. FEBS Letters. 487 (2), 203-208 (2000).
- Firer, M. A. Efficient elution of functional proteins in affinity chromatography. Journal of Biochemical and Biophysical Methods. 49 (1-3), 433-442 (2001).
- Pabst, T. M., et al. Engineering of novel Staphylococcal Protein A ligands to enable milder elution pH and high dynamic binding capacity. Journal of Chromatography A. 1362, 180-185 (2014).
- Müller, E., Vajda, J. Routes to improve binding capacities of affinity resins demonstrated for Protein A chromatography. Journal of Chromatography B. 1021, 159-168 (2016).
- Shamloul, M., Trusa, J., Vadim, M., Vidadi, Y. Optimization and Utilization of Agrobacterium-mediated Transient Protein Production in Nicotiana. Journal of Visualized Experiments. (86), 51204 (2014).
- Rademacher, T., et al. Plant cell packs: a scalable platform for recombinant protein production and metabolic engineering. Plant Biotechnology Journal. , (2018).
- Saxena, L., Iyer, B. K., Ananthanarayan, L. Purification of a bifunctional amylase/protease inhibitor from ragi (Eleusine coracana) by chromatography and its use as an affinity ligand. Journal of Chromatography. B. 878 (19), 1549-1554 (2010).
- Kurppa, K., Reuter, L. J., Ritala, A., Linder, M. B., Joensuu, J. J. In-solution antibody harvesting with a plant-produced hydrophobin-Protein A fusion. Plant Biotechnology Journal. 16 (2), 404-414 (2018).
- Menzel, S., et al. Optimized Blanching Reduces the Host Cell Protein Content and Substantially Enhances the Recovery and Stability of Two Plant-Derived Malaria Vaccine Candidates. Frontiers in Plant Science. 7 (159), 1-7 (2016).
- Yigzaw, Y., Piper, R., Tran, M., Shukla, A. A. Exploitation of the adsorptive properties of depth filters for host cell protein removal during monoclonal antibody purification. Biotechnology Progress. 22 (1), 288-296 (2006).
- Menzel, S., et al. Downstream processing of a plant-derived malaria transmission-blocking vaccine candidate. Protein Expression and Purification. 152, 122-130 (2018).
- Vöpel, N., Boes, A., Edgü, G., Beiss, V. Malaria vaccine candidate antigen targeting the pre-erythrocytic stage of Plasmodium falciparum produced at high level in plants. Biotechnology Journal. 9 (11), 1435-1445 (2014).
- Lee, C. W., Wang, H. J., Hwang, J. K., Tseng, C. P. Protein thermal stability enhancement by designing salt bridges: a combined computational and experimental study. PLoS ONE. 9 (11), e112751 (2014).
- Zhu, L., Cheng, F., Piatkowski, V., Schwaneberg, U. Protein engineering of the antitumor enzyme PpADI for improved thermal resistance. Chembiochem. 15 (2), 276-283 (2014).
- Khanal, O., et al. Contributions of depth filter components to protein adsorption in bioprocessing. Biotechnology and Bioengineering. 115 (8), 1938-1948 (2018).
- Buyel, J. F., Fischer, R. Downstream processing of biopharmaceutical proteins produced in plants: the pros and cons of flocculants. Bioengineered. 5 (2), 138-142 (2014).
- Buyel, J. F. Procedure to Evaluate the Efficiency of Flocculants for the Removal of Dispersed Particles from Plant Extracts. Journal of Visualized Experiments. (110), e53940 (2016).
- Fink, D., et al. Ubiquitous expression of the monomeric red fluorescent protein mCherry in transgenic mice. Genesis. 48 (12), 723-729 (2010).
- Gagnon, P. Technology trends in antibody purification. Journal of Chromatography A. 1221, 57-70 (2012).
- Figueira, M., Laramée, L., Murrell, J. C., Groleau, D., Míguez, C. Production of green fluorescent protein by the methylotrophic bacterium Methylobacterium extorquens. FEMS Microbiology Letters. 193 (2), 195-200 (2001).
- Bornhorst, J. A., Falke, J. J. Purification of proteins using polyhistidine affinity tags. Methods in enzymology. 326, 245-254 (2000).
- Sainsbury, F., Jutras, P. V., Vorster, J., Goulet, M., Michaud, D. A. Chimeric Affinity Tag for Efficient Expression and Chromatographic Purification of Heterologous Proteins from Plants. Frontiers in Plant Science. 7, 141-141 (2016).
- Krupka, M., et al. The Position of His-Tag in Recombinant OspC and Application of Various Adjuvants Affects the Intensity and Quality of Specific Antibody Response after Immunization of Experimental Mice. PLoS ONE. 11 (2), e0148497 (2016).
- Goel, A., et al. Relative position of the hexahistidine tag effects binding properties of a tumor-associated single-chain Fv construct. Biochimica et Biophysica Acta. 1523 (1), 13-20 (2000).
- Tustian, A. D., et al. Development of a novel affinity chromatography resin for platform purification of bispecific antibodies with modified protein a binding avidity. Biotechnology Progress. , (2018).
- Zandian, M., Jungbauer, A. An immunoaffinity column with a monoclonal antibody as ligand for human follicle stimulating hormone. Journal of Separation Science. 32 (10), 1585-1591 (2009).
- Kelley, B. Very large scale monoclonal antibody purification: the case for conventional unit operations. Biotechnology Progress. 23 (5), 995-1008 (2007).
- Petrou, C., Sarigiannis, Y., S, K. o. u. t. s. o. p. o. u. l. o. s. Ch. 1. Peptide Applications in Biomedicine, Biotechnology and Bioengineering. , 1-21 (2018).
- Menegatti, S., et al. Design of protease-resistant peptide ligands for the purification of antibodies from human plasma. Journal of Chromatography A. 1445, 93-104 (2016).
- Rademacher, T., et al. Plant cell packs: a scalable platform for recombinant protein production and metabolic engineering. Plant Biotechnology Journal. , 1-7 (2019).
