I betragtning af søvnens udbredelse i vores daglige rutine, er det vigtigt at forstå dens funktion. Undersøgelser med dette mål kræver præcis måling af søvn. Polysomnografi (PSG) er den gyldne standard måling af søvn. PSG giver mulighed for objektiv, kvantitativ måling af søvn med høj tidsmæssig opløsning og kan være nyttig til både forskning og kliniske formål. PSG er en kombination af fysiologiske optagelser. Som minimum omfatter en PSG montage følgende foranstaltninger: elektroencefalografi (EEG), elektrooculografi (EOG) og Elektromyografi (EMG). Disse foranstaltninger vurderer elektriske potentialer fra henholdsvis hjernen, øjnene og musklerne og giver mulighed for klassifikation af søvnstadier (Se figur 1). Andre foranstaltninger, såsom elektro kardiografi (EKG), respiration, og pulsoximetri kan inkluderes for at identificere tilstedeværelsen af uordnet søvn.
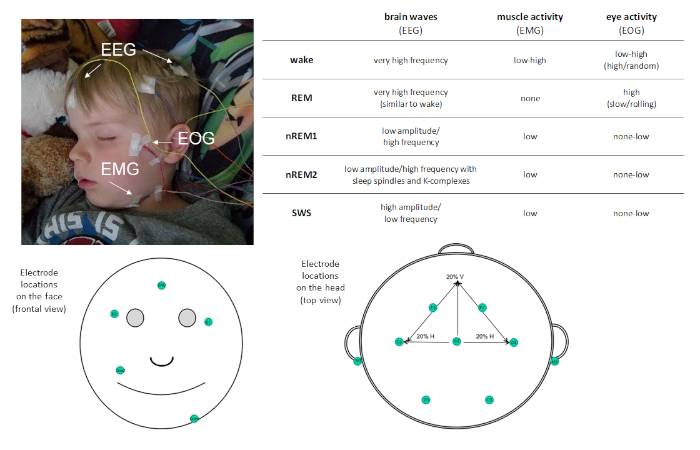
Figur 1: eksempel på elektrodeplacering og beskrivelse af aktivitet registreret via PSG. Venligst klik her for at se en større version af dette tal.
PSG tillader søvn at være karakteriseret i fire forskellige søvnstadier: ikke-hurtig øjenbevægelse (ikke-REM) etape 1 (nREM1; 4 − 7 Hz), ikke-REM fase 2 (nREM2; 12 − 15 Hz), og ikke-REM fase 3 (mere almindeligt kendt som Slow Wave Sleep [SWS]; 0,5 − 4 Hz), og Rapid-Eye bevægelse (REM søvn. nREM1 markerer søvn debut, og er identificeret baseret på reduceret muskeltonus i EMG omkodning og blandede amplitude EEG svingninger i forhold til Alpha observeret i hvile kølvandet. Dette efterfølges af nREM2, som kan skelnes ved tilstedeværelsen af Sleep spindler (korte byger af Sigma frekvens aktivitet; 11 − 16 Hz) og K-komplekser (enkelt langsom-bølger, der skiller sig ud fra den omgivende aktivitet) i EEG. SWS er karakteriseret ved distinkte, langsomme EEG-svingninger med høj amplitude. REM søvn er karakteriseret ved hurtig lav-amplitude oscillerende hjerneaktivitet meget lig den observerede under kølvandet. Men det, der skelner REM-søvn fra Wake er, at det også er karakteriseret ved phasic Rapid Eye bevægelser (deraf moniker REM) og muskel atonia. I løbet af en søvn bout, søvnstadier opleves cyklisk, med en hastighed på omkring 90 min/cyklus.
Søvn følger også døgnrytmen, med søvnanfald, der finder sted i 24-h cyklusser. Søvn timing og konsistens kan påvirke søvn funktion og er også vigtigt at vurdere. Selvom PSG er nødvendig for at karakterisere søvnstadier, er det tidskrævende at anvende og derfor ikke ideel til vurdering af flere søvnanfald (f. eks. flere overnatninger med søvn, lur og overnatning). Til dette, actigraphy er gavnlig. Actigraphy bruger et Tri-Axial accelerometer, typisk på håndleddet, til at estimere søvn baseret på fravær af bevægelse. Selvom actigrafi ikke kan bruges til at karakterisere søvnstadier, det har vist sig at være pålidelige til påvisning søvn debut og vågne debut (herunder søvn fragmentering eller vågne efter søvn debut) i en række populationer fra spædbørn1 til ældre voksne2 . Både PSG og actigraphy er foretrukne metoder over selv/forælder-rapport foranstaltninger. Selv/forælder-rapport foranstaltninger er nemme at administrere og relativt billig, men de er også underlagt bias og manglende overholdelse. Endelig, det er værd at bemærke, at disse metoder kan bruges i kombination til at udnytte de stærke sider af hver. For eksempel kan PSG kombineres med actigrafi og/eller selv/forælder-rapport for at opnå både overnatning søvnkvalitet samt verifikation af søvn mængder eller søvn-vågne cyklusser, især over lange varigheder (f. eks uger eller måneder).
En funktion af søvn, der har høstet særlig interesse er søvn afhængig hukommelse konsolidering, behandling af erindringer, der efterlader dem stærkere og mindre sårbar over for indblanding3. Selvom hukommelses konsolidering kan finde sted under Wake in Children4 og Adults5, er der betydelige beviser for, at konsolideringen forstærkes under søvnen. Tidligere forskning har givet adfærdsmæssige eksempler på søvn afhængig hukommelse konsolidering ved at sammenligne ændringer i hukommelsen ydeevne efter et interval af søvn (f. eks 8 PM − 8 am) til ændringer efter et tilsvarende interval brugt vågen (f. eks 8 am − 8 PM). Hos voksne er minderne beskyttet6 eller endda forbedret7 efter et interval af søvn, mens minderne typisk henfalder over et tilsvarende interval af Wake. Der er anvendt kontrol, som dissocierer præstations ændringer fra cirkadiske påvirkninger8,9,10. For eksempel observeres lignende fordele ved søvn, når du sammenligner præstationer over en Mid-Day NAP til en tilsvarende Mid-Day Wake periode9.
Selv om søvn var engang menes at afspejle en passiv proces, blot beskytte erindringer fra forfald eller indblanding, moderne teorier tyder søvn spiller en mere aktiv rolle og faktisk fremmer hukommelse gennem reaktiveringer11,12 ,13. Støtte til dette kommer fra observerede korrelationer mellem adfærdsmæssige foranstaltninger af hukommelse konsolidering over søvn (ændring i hukommelsen tilbagekaldelse efter søvn i forhold til før søvn) og specifikke aspekter af søvn fysiologi. For mange deklarative hukommelses opgaver er hukommelses konsolidering forbundet med aspekter af ikke-REM-søvn, specifikt målinger af SWS eller Sleep-spindler, der findes i nREM2 og SWS. Hvis søvnens rolle var passiv beskyttelse mod interferens, ville en sådan sammenhæng ikke forventes; snarere en sammenhæng mellem tid i søvn (uanset søvn fase) og ydeevne ville forventes, som mere tid i søvn ville give mere beskyttelse mod indblanding14.
Yderligere støtte til den aktive rolle SWS i hukommelsen konsolidering er tydelig i undersøgelser af målrettede hukommelse reaktivering. I disse undersøgelser, en hukommelse er lært i forbindelse med en perceptuelle cue, for eksempel en lugt, og tilbagekaldelse af hukommelsen er større efter søvn, hvis cue er re-præsenteret under søvn, SWS i særdeleshed15. Selv om den underliggende mekanisme er drøftet16,17, en fremtrædende teori, systemer konsolidering teori, hævder, at erindringer kodet i hippocampus er stabiliseret i cortex selv hippocampal-neocortical dialog. Specifikt, kortikale langsomme bølger og sove spindler, der opstår i forbindelse med hippocampus krusninger forbundet med hukommelse reaktivering, støtte hukommelsen overførsel3.
Den rolle søvn i hukommelsen konsolidering under udvikling er mindre klar. Tidlig barndom er en periode af særlig interesse som børn begynder at overgå fra en bifasisk (bestående af en Mid-Day lur og en overnatning søvn bout) til en monofasiske søvnmønster. Nyere forskning tyder på, at denne overgang kan afspejle hjerne modning18. Dette argument er i overensstemmelse med empiriske data viser udviklingsmæssige ændringer i natten søvn (dvs. topografi af langsom bølge aktivitet) spejle, at af kortikale modning19.
Selv om der er flere adfærdsmæssige demonstrationer af natten søvn-afhængige konsolidering i børn20,21 og spædbørn22, forskning på neurale funda-ter af hukommelse konsolidering med Mid-dag søvn er lige begyndt at blive undersøgt. I banebrydende arbejde undersøger rollen som Mid-Day nationale handlingsplaner på hukommelse i førskolebørn, nationale handlingsplaner viste sig at beskytte erindringer af nyligt lærte oplysninger, mens hukommelsen blev reduceret (ved ~ 12%) da børn forblev vågne under NAP-intervallet23. Denne “NAP benefit” var størst hos børn, der misligholdte sædvanligvis (dvs. 5 eller flere gange om ugen målt med actigrafi) uanset deres alder. Ved at optage PSG i løbet af NAP blev det konstateret, at ændringen i hukommelsens ydeevne på tværs af NAP-perioden var specifikt forbundet med søvn spindel tæthed (antallet af Sleep-spindler pr. minut af nREM), hvilket tyder på NAP-kvalitet (ikke kvantitet) var en kritisk faktor i fremme af hukommelses fastholdelse (Se afsnittet om repræsentative resultater).
Denne undersøgelse fremhæver betydningen af PSG i at udforske forholdet mellem søvn og hukommelse under udvikling. Det peger på vigtigheden af at karakterisere søvn makro-(søvnstadier) og mikro-(kvaliteter af disse stadier såsom frekvenser og tilstedeværelsen af spindler) strukturer under lur for hukommelse konsolidering. Det fremhæver også vigtigheden af at vurdere søvn rytmer (karakterisering af børn som sædvanligt eller ikke-sædvanligt serviet). Selv om vores arbejde har karakteriseret funktionen af nationale handlingsplaner i visuospatiale læring (og mere for nylig følelsesmæssige24 og procedure25 læring), mange spørgsmål tilbage. For eksempel vil det være vigtigt at undersøge andre deklarative hukommelse opgaver for at vurdere generalisering af disse resultater og til at vurdere opgaver, der anvendes i førskole klasseværelser til at forstå specifikke parametre (f. eks, mængden af NAP fordel i forhold til læring) for miljømæssigt gyldige opgaver. Yderligere arbejde vil også være nødvendigt at forstå, når Wake er tilstrækkelig til hukommelse konsolidering. Således er vores mål at afystificere processen med at måle søvn og søvn afhængig hukommelse konsolidering hos børn. Vi giver praktiske tips til at undersøge fordelen ved en eftermiddagslur på deklarativ hukommelse i typisk at udvikle førskolebørn (ca. 3 til 4 år) ved hjælp af en edb visuospatiale hukommelse opgave samt metoder til vurdering af NAP habituality bruge actigraphy, Parent-Report og NAP fysiologi ved hjælp af PSG. Selv om disse metoder blev udviklet til førskolebørn, der NAP med varierende hyppighed, disse metoder kan tilpasses til enhver aldersgruppe.
