Le cancer est l’une des principales causes de décès dans le monde. Par conséquent, il y a une demande pour développer des modèles expérimentaux fiables de maladie pour obtenir une meilleure compréhension de la maladie aussi bien que pour explorer des approches thérapeutiques potentielles. L’un des modèles expérimentaux in vivo les plus couramment utilisés pour étudier le développement du cancer de la peau est le modèle de carcinogenèse de la peau à deux étapes induit chimiquement à deux étapes1,2. Le modèle fournit un outil pour étudier l’initiation, la promotion et la progression de tumeur en plus des événements spécifiques tels que l’infiltration de cellules immunitaires et l’angiogenèse.
Pour employer le modèle de carcinogenèse de peau de deux étapes, la peau arrière des souris est traitée avec deux produits chimiques différents qui induisonnt ensemble la formation de tumeur. Le modèle est initié avec une faible dose de la mutagène, DMBA, suivie d’une exposition prolongée au promoteur de la tumeur, TPA3 (Figure 1). DMBA mute l’ADN au hasard en formant des adducts covalents avec l’ADN des cellules épidermiques et des cellules souches de kératinocyte primaire4,5,6,7. Certaines de ces mutations aléatoires ont lieu dans un proto-oncogène, comme Hras1 (mutations dans Kras et Nras sont également détectées) et la conversion de proto-oncogènes en oncogènes conduit la formation tumorale sous des stimuli appropriés. TPA, à son tour, est l’agent de croissance de croissance de tumeur le plus couramment utilisé. Sa cible moléculaire est la protéine kinase C (PKC)8. TPA active également la signalisation Wnt/ -caténine qui est cruciale pour la formation de tumeur dans le modèle9. L’exposition répétée et prolongée à l’agent de promotion mène à la signalisation accrue de cellules, à la production accrue des facteurs de croissance, et à une réaction inflammatoire locale, qui sont évidentes dues à la synthèse accrue d’ADN et à l’infiltration inflammatoire de cellules dans la peau traitée.
Les principaux médiateurs inflammatoires du modèle DMBA-TPA ont été identifiés10. Interleukin-17A (IL-17A) est connu pour être particulièrement tumorigène dans le modèle DMBA-TPA11,12. Il travaille en synergie avec l’interleukine 6 (IL-6) et participe au recrutement de macrophage et de neutrophile13,14. En outre, les cellules CD4et T et les neutrophiles se sont avérés tumorigènes dans le modèle DMBA-TPA. Enfin, les macrophages peuvent également promouvoir la tumorigénèse dans le modèle15,16,17.
Pendant la phase de promotion, la prolifération cellulaire des cellules mutées est améliorée et une hyperplasie soutenue de l’épiderme est maintenue1. Cela conduit au développement du papillome dans la peau en 10-20 semaines, après quoi les papillomes commencent à se convertir en tumeurs malignes, carcinomes épidermoïdes (SCC)2. Cependant, moins de 10% des papillomes progressent vers la malignité, bien que ce pourcentage dépende également du fond génétique des souris2,18. Pendant des décennies, on ne savait pas quel type de cellules ont été initialement mutés dans les tumeurs conduisant à la malignité, même si certaines études avaient rapporté des caractéristiques clairement distinctes dans les tumeurs malignes par rapport aux papillomes bénins19,20. Cependant, des études récentes ont considérablement augmenté notre compréhension sur l’origine clonale de la formation de tumeur dans le modèle21de DMBA-TPA. 22. 23. Il a été démontré que les deux cellules épithéliales dérivées de moelle osseuse et les cellules souches pileux follicules contribuent à la formation de tumeur22. Des études de traçage de lignée scénographie spécifiques à la scène ont révélé que les papillomes bénins sont d’origine monoclonal, mais ils recrutent de nouvelles populations épithéliales21,23. Cependant, un seul des clones cellulaires fonctionne comme un moteur de la carcinogenèse; il contient une mutation hras23. La progression vers la formation de carcinome est associée à un balayage clonal23.
Le DMBA cancérogène initie la formation de papillome et TPA favorise la croissance tumorale. Par conséquent, l’initiation de tumeur peut être étudiée séparément de la promotion en interrompant l’expérience avant la période de traitement de TPA. Comme la progression de tumeur est étudiée hebdomadairement elle offre une grande occasion pour l’analyse détaillée de croissance de tumeur tout au long de l’étude. Puisque les tumeurs sont produites par des produits chimiques externes, une mutation oncogène dans la lignée germinale est inutile. Ainsi, l’étude des effets d’un fond génétique (par exemple, knock-out/transgène contre type sauvage) sur la tumorigénèse est simple2. En somme, le modèle de cancer de la peau DMBA/TPA est une approche particulièrement utile pour étudier le rôle du système immunitaire dans la progression tumorale ainsi que pour l’évaluation des étapes d’initiation et de promotion de tumeur indépendamment ou interdépendante.
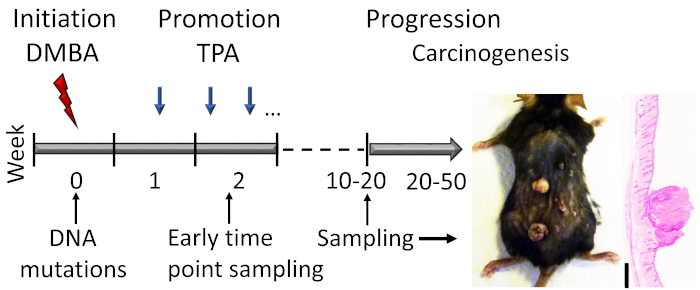
Figure 1 : Contour du modèle de carcinogenèse de la peau induit par DMBA-TPA. Le DMBA cancérogène est appliqué topiquement pour induire des mutations d’ADN dans la phase d’initiation du modèle. L’agent de promotion de la croissance TPA est administré 2x par semaine pour améliorer la prolifération cellulaire pendant la phase de promotion, conduisant au développement de papillomes dans la peau. Les animaux sont sacrifiés après que la réponse du papillome a atteint un plateau, généralement dans les semaines 15-20, selon le fond génétique des souris. Une petite proportion des papillomes peut se développer davantage en SCC dans un délai de 20 à 50 semaines. Pour étudier les premiers événements de la phase d’initiation et de promotion précoce, des échantillons peuvent être prélevés (p. ex., peu de temps après la deuxième demande d’APT). Une photographie représentative et l’hématoxylin et l’éosine souillées section transversale des papillomes sur une peau de souris C57BL/6 après 19 semaines de traitement sont montrées. Barre d’échelle de 0,1 mm. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
