कैंसर दुनिया में मौत के प्रमुख कारणों में से एक है। इसलिए, रोग की बेहतर समझ प्राप्त करने के साथ-साथ संभावित चिकित्सीय दृष्टिकोणों का पता लगाने के लिए विश्वसनीय प्रयोगात्मक रोग मॉडल विकसित करने की मांग की गई है। त्वचा कैंसर के विकास का अध्ययन करने के लिए वीवो मॉडल में सबसे अधिक उपयोग किए जाने वाले प्रयोगात्मक में से एक रासायनिक रूप से प्रेरित दो-चरण त्वचा कार्सिनोजेनेसिस मॉडल1,2है। मॉडल प्रतिरक्षा सेल घुसपैठ और एंजियोजेनेसिस जैसी विशिष्ट घटनाओं के अलावा ट्यूमर दीक्षा, पदोन्नति और प्रगति का अध्ययन करने के लिए एक उपकरण प्रदान करता है।
दो चरण त्वचा कार्सिनोजेनेसिस मॉडल का उपयोग करने के लिए, चूहों की पीठ की त्वचा को दो अलग-अलग रसायनों के साथ इलाज किया जाता है जो एक साथ ट्यूमर गठन को प्रेरित करते हैं। मॉडल को म्यूटाजेन, DMBA की कम खुराक के साथ शुरू किया जाता है, जिसके बाद ट्यूमर प्रमोटर, टीपीए3 (चित्रा 1)के लिए लंबे समय तक एक्सपोजर होता है। डीएमबीए एपिडरमल कोशिकाओं और प्राथमिक केराटिनोसाइट स्टेम सेल4,5,6,7के डीएनए के साथ सहसंयोजक एडक्ट्स बनाकर डीएनए को बेतरतीब ढंग से म्यूट करता है । इनमें से कुछ यादृच्छिक उत्परिवर्तन एक प्रोटो-ऑन्कोजीन में होते हैं, जैसे कि Hras1 (Kras और Nras में उत्परिवर्तन भी पता लगाया जाता है) और प्रोटो-ऑन्कोजीन का ऑन्कोजीन में रूपांतरण उचित उत्तेजनाओं के तहत ट्यूमर गठन को चलाता है। टीपीए, बदले में, सबसे अधिक इस्तेमाल किया ट्यूमर विकास को बढ़ावा देने एजेंट है । इसका आणविक लक्ष्य प्रोटीन किनाज सी (पीकेसी)8है । टीपीए भी Wnt/-catenin संकेत है कि मॉडल9में ट्यूमर गठन के लिए महत्वपूर्ण है सक्रिय करता है । को बढ़ावा देने वाले एजेंट के लिए दोहराया और लंबे समय तक जोखिम बढ़ाया सेल संकेत, विकास कारकों के उत्पादन में वृद्धि, और एक स्थानीय भड़काऊ प्रतिक्रिया है, जो डीएनए संश्लेषण और इलाज त्वचा में भड़काऊ सेल घुसपैठ के कारण स्पष्ट कर रहे है की ओर जाता है ।
DMBA-टीपीए मॉडल में प्रमुख भड़काऊ मध्यस्थों की पहचान10की गई है । इंटरल्यूकिन-17ए (आईएल-17ए) को DMBA-टीपीए मॉडल11,12में विशेष रूप से ट्यूमरीजेनिक माना जाता है। यह इंटरल्यूकिन 6 (आईएल-6) के साथ तालमेल में काम करता है और मैक्रोफेज और न्यूट्रोफिल भर्ती13,14में भाग लेता है । इसके अलावा डीएमबीए-टीपीए मॉडल मेंसीडी4+ टी सेल और न्यूट्रोफिल को ट्यूमरीजेनिक दिखाया गया है। अंत में, मैक्रोफेज मॉडल15,16,17में ट्यूमरजेनेसिस को भी बढ़ावा दे सकता है।
पदोन्नति चरण के दौरान, उत्परिवर्तित कोशिकाओं के कोशिका प्रसार को बढ़ाया जाता है और एपिडर्मिस की निरंतर हाइपरप्लासिया1बनाए रखी जाती है। इससे त्वचा में 10-20 सप्ताह में पेपिलोमा विकास होता है, जिसके बाद पेपिलोमा घातक ट्यूमर, स्क्वैमस सेल कार्सिनोमा (एससीसी)2में परिवर्तित होना शुरू हो जाता है। हालांकि, पेपिलोमा का 10% से भी कम द्रोह करने के लिए प्रगति करता है, हालांकि यह प्रतिशत चूहों2,18की आनुवंशिक पृष्ठभूमि पर भी निर्भर करता है। दशकों से यह ज्ञात नहीं था कि ट्यूमर में शुरू में किस प्रकार की कोशिकाओं को द्रोह की ओर ले जाया गया था, भले ही कुछ अध्ययनों ने सौम्य पेपिलोमास19,20की तुलना में घातक ट्यूमर में स्पष्ट रूप से अलग विशेषताओं की सूचना दी थी। हालांकि, हाल के अध्ययनों से DMBA-टीपीए मॉडल21में ट्यूमर गठन की क्लोनल उत्पत्ति पर हमारी समझ बहुत बढ़ गईहै। 22. 23. यह प्रदर्शन किया गया था कि दोनों अस्थि मज्जा व्युत्पन्न एपिथेलियल कोशिकाओं और बाल कूप स्टेम कोशिकाओं ट्यूमर गठन22में योगदान . स्टेज-विशिष्ट वंश ट्रेसिंग अध्ययनों का अनावरण किया गया है कि सौम्य पेपिलोमा मोनोक्लोनल मूल के हैं, लेकिन वे21,23नए एपिथेलियल सेल आबादी की भर्ती करते हैं। हालांकि, केवल एक सेल क्लोन कार्सिनोजेनेसिस के लिए एक ड्राइवर के रूप में कार्य करता है; यह एक Hras म्यूटेशन23शामिल हैं . कार्सिनोमा गठन की प्रगति क्लोनल स्वीप23से जुड़ी हुई है ।
कार्सिनोजन DMBA पेपिलोमा गठन शुरू करता है और टीपीए ट्यूमर के विकास को बढ़ावा देता है। इसलिए, टीपीए उपचार अवधि से पहले प्रयोग में व्यवधान डालकर ट्यूमर दीक्षा का प्रचार से अलग से अध्ययन किया जा सकता है। के रूप में ट्यूमर प्रगति साप्ताहिक अध्ययन किया है यह अध्ययन के दौरान विस्तृत ट्यूमर विकास विश्लेषण के लिए एक महान अवसर प्रदान करता है । क्योंकि ट्यूमर बाहरी रसायनों से उत्पन्न होते हैं, अंकुरित में एक ऑन्कोजेनिक उत्परिवर्तन अनावश्यक है। इस प्रकार, ट्यूमरजीनिस पर आनुवंशिक पृष्ठभूमि (जैसे, नॉकआउट/ट्रांसजीन बनाम जंगली प्रकार) के प्रभावों का अध्ययन करना सीधा2है । संक्षेप में, DMBA/टीपीए त्वचा कैंसर मॉडल ट्यूमर प्रगति में प्रतिरक्षा प्रणाली की भूमिका का अध्ययन करने के साथ ही ट्यूमर दीक्षा और संवर्धन कदम स्वतंत्र रूप से या परस्पर निर्भर के मूल्यांकन के लिए एक विशेष रूप से उपयोगी दृष्टिकोण है ।
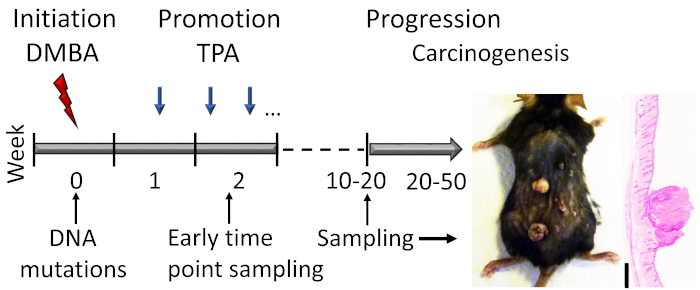
चित्रा 1: DMBA-टीपीए प्रेरित त्वचा कैंसरजन्य मॉडल रूपरेखा। कार्सिनोजन DMBA मॉडल के दीक्षा चरण में डीएनए म्यूटेशन को प्रेरित करने के लिए सामयिक रूप से लागू किया जाता है। विकास को बढ़ावा देने वाले एजेंट टीपीए को पदोन्नति चरण के दौरान सेल प्रसार को बढ़ाने के लिए एक सप्ताह में 2x प्रशासित किया जाता है, जिससे त्वचा में पेपिलोमा का विकास होता है। जानवरों की बलि के बाद papilloma प्रतिक्रिया एक पठार तक पहुंचता है, आमतौर पर सप्ताह के भीतर 15-20, चूहों की आनुवंशिक पृष्ठभूमि पर निर्भर करता है । पेपिलोमा का एक छोटा सा अनुपात 20-50 सप्ताह के भीतर एससीसी में और विकसित हो सकता है। दीक्षा और प्रारंभिक पदोन्नति चरण में प्रारंभिक घटनाओं का अध्ययन करने के लिए, नमूने एकत्र किए जा सकते हैं (उदाहरण के लिए, दूसरे टीपीए आवेदन के तुरंत बाद)। उपचार के 19 सप्ताह के बाद C57BL/6 माउस त्वचा पर पेपिलोमास के एक प्रतिनिधि तस्वीर और हेमैटोक्सिलिन और eosin दाग पार अनुभाग दिखाया जाता है । स्केल बार = 0.1 मिमी। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
