The current modified protocol allows efficient isolation and culture of rat and mice CMs in vitro. For rat CM isolation, a total of 3 adult (12-week-old) male Fischer 344 rats were used in the procedure. Figure 1 shows the surgical apparatus and isolation setups that are required in the procedure; each part has been marked and described in the figure legend. Collagenase type 2 was used for digestion, which yields a high quantity of high quality CMs from successful isolation (Figure 2A). Twenty-four hours post-isolation, these cells were transfected with cell cycle inducing specific siRNAs against Rb1 and Meis2, whereas, cel-miR-67 was used as transfection control in the experiment11. CMs were maintained in culture for either seven days (Figure 2B) or up to twenty days (Figure 2C) to observe the morphological changes. To score for cell cycle activity, CMs were fixed on day 7 and stained for cardiac-specific marker Troponin-I, mitotic marker KI67, and nucleus was visualized through DAPI (Figure 2D-F). A significant increase in KI67 positive cardiomyocytes was observed with siRb1+siMeis2 treatment when compared to control11. Moreover, the rat cardiomyocytes were beating (contractile function) in culture on day seven post-plating, which is a hallmark characteristic of healthy cardiomyocytes11.
For CM isolation in mice, a total of 3 male and 3 female adult (12-week-old) C57BL/6 mice were used in the procedure. Similar to the rat study, collagenase type 2 was used to digest the collagen and the extracellular matrix of the adult mice heart. Adult mice CM is comparatively fragile for isolation and culture in vitro; thus, this protocol uses blebbistatin to improve the viability of the adult mouse CM. Successful isolation yields >70-80% healthy rod shape CMs, which can be cultured for up to 20 days (Figure 3A-C) and maintain their cardiac Troponin-I staining and contractility (Figure 3C). In separate experiments, four hours post isolation, mouse CMs were transfected with a cell cycle inducing siRNA cocktail as described for the rat CM. Briefly, mice CM was simultaneously transfected with the specific siRNAs against Rb1 and Meis2, and control (cel-miR-67). Post-transfection CMs were subject to time course imaging for 10 days showing the morphological changes (Figure 4) which is followed by a proliferation assay (Figure 5). On day 10, immune-fluorescent staining for Troponin-I, KI67, and DAPI was performed as previously described. A significant increase in KI67 positive cardiomyocytes was observed with siRb1+siMeis2 treatment when compared to control (Figure 5A-C).
The present protocol demonstrates that adult rats and mice CMs can be cultured long-term in vitro for up to 20 days and potentially longer. After 20 days, CMs maintain their Troponin-I staining as well as contractility if the inhibitors are removed from the media.
| Myocyte buffer composition, pH 7.4 (Prepare and can be stored at 4 °C ) | |||
| Reagent | Concentration, mM | Molecular wt. | For 1 Liter |
| NaCl | 113 | 58.44 | 6.6037 g |
| KCl | 4.7 | 74.5513 | 350.3911 mg |
| MgSO4 | 1.2 | 120.366 | 144.4392 mg |
| KH2PO4 | 0.6 | 136.086 | 81.6516 mg |
| NaH2PO4 | 0.6 | 119.98 | 71.988 mg |
| Perfusion buffer, pH 7.4 (Prepare fresh for each isolation) | |||
| Concentration, mM | Molecular wt. | Amount required | |
| Myocyte buffer | 250 mL | ||
| HEPES | 10 | 238.3012 | 595.75 mg |
| 2,3-butanedione monoximine | 10 | 101.105 | 252.775 mg |
| NaHCO3 Fresh | 1.6 | 84.007 | 33.6028 mg |
| Taurine Fresh | 30 | 125.15 | 938.625 mg |
| Glucose Fresh | 20 | 180.156 | 900.775 mg |
| Enzyme solution | |||
| Stock Conc. | Working Conc. | Required | |
| Myocyte buffer | 50 mL | 50 mL | |
| Collagenase type 2 | 330 U/mg | 620 U/mL | 93 mg |
| Protease XIV | >3.5 U/mg | 0.104 U/mL | 1.48 mg |
| DNase I grade 2 | 0.015 mg/mL | ||
| Stop Buffer A | |||
| Stock Conc. | Working Conc. | Required | |
| Myocyte buffer | 30 mL | ||
| BSA | 2.50% | 0.75 g | |
| CaCl2 | 100 mM | 0.1 mM | 30 µL |
| Stop Buffer B | |||
| Stock Conc. | Working Conc. | Required | |
| Myocyte buffer | 30 mL | ||
| BSA | 5.00% | 1.5 g | |
| CaCl2 | 100 mM | 0.1 mM | 30 µL |
Table 1: Solutions required for adult mouse cardiomyocyte isolation.
| 10x KHB Stock Solution (Total volume= 1L) |
Reagent | Molarity (mM) | Amount (g) |
| NaCl | 1180 | 68.9 g | |
| KCl | 48 | 3.5 g | |
| HEPES | 250 | 59.7 g | |
| MgSO4 | 12.5 | 1.4 g | |
| K2HPO4 | 12.5 | 2.1 g | |
| Adjust pH to 7.4 with 4 M NaOH (~20 mL), store at 4 °C | |||
| KHB Solution, 500 mL | Reagent | Amount | |
| 10x KHB | 50 mL | ||
| Glucose | 0.99 g | ||
| Taurine | 0.31 g | ||
| Add H2O to bring volume to 500 mL, and pH should be ~7.35 | |||
| Solution A | Reagent | Amount | |
| KHB solution | 500 mL (10 mM) | ||
| BDM | 0.5 g | ||
| Oxygenate with 100% O2 and warm to 37 °C | |||
| Solution B, 50 mL | Reagent | Amount | |
| Solution A | 50 mL | ||
| BSA | 0.5 g | ||
| 0.1 M CaCl2 (Ca++=0.1 mM) | 50 µL | ||
| Solution E, 50mL | Reagent | Amount | |
| Solution A | 50 mL | ||
| BSA | 0.05 g | ||
| Collagenase type II (263 units/mg) | 35 mg | ||
| Hyaluronidase (Type I-S) | 10 mg | ||
| 0.1 M CaCl2 stock | 12.5 µL | ||
| Mix well | |||
| CaCl2 Stock, 0.1M | Reagent | Amount | |
| CaCl2 | 7.35 g | ||
| H2O | 500 mL | ||
| Store at 4 °C | |||
Table 2: Solutions required for adult rat cardiomyocyte isolation.
| Plating media composition, pH 7.4 | |||
| Working Concentration | Molecular Wt. | Required amount | |
| Culture media without Blebbistatin | 50 mL | ||
| BDM | 10 mM | 101.105 g/mol | 50.55 mg |
| FBS | 5% | 2.5 g | |
| Culture media composition, pH 7.4 | |||
| Reagent | Working Concentration | Molecular Wt. | Required amount |
| DMEM | 1X | 250 mL | |
| Insulin | 1 µg/mL | 0.25 mg | |
| Transferrin | 0.55 µg/mL | 0.138 mg | |
| Selenium | 0.5 ng/mL | 0.125 µg | |
| Penicillin (U/mL)-streptomycin (g/mL) | 100-100 | 2.5 mL | |
| HEPES | 10 mM | 238.3012 g/mol | 595.753 mg |
| *FBS | 10% | 25 mL | |
| *BSA | 0.20% | 0.5 g | |
| #Blebbistatin | 25 µM | 292.338 g/mol | 1.8271 mg |
| *Use either of them, as per the experimental requirement. # Aliquot the culture media to prepare plating media before adding Blebbistatin. NOTE: Prepare 200 mL of culture media. NOTE: Prepare 50 mL of plating media. |
|||
Table 3: Media composition for adult mouse cardiomyocyte plating and culture.
| Culture media composition, pH 7.4 | ||
| Reagent | Working Concentration | Required amount |
| DMEM | 1x | 250 mL |
| Penicillin (U/mL)-streptomycin (g/mL) | 100-100 | 2.5 mL |
| *FBS | 10% | 25 mL |
Table 4: Media composition for adult rat cardiomyocyte plating and culture.
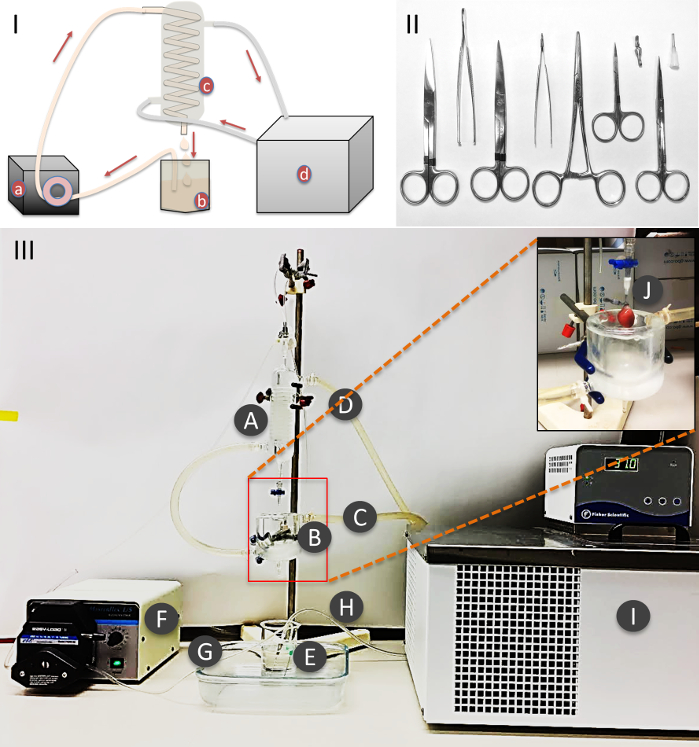
Figure 1: Procedure setup and equipment. (I) Schematic representation of the perfusion. (II) Surgical instruments and cannulation needle. (III) Heart perfusion assembly: A) Heating jacket. B) Double wall water jacket vessel. C) Circulating heated water inlet. D) Circulating heated water outlet. E) Heart perfusion solution. F) Circulating pump G) Perfusion solution tube. H) Oxygen supply tube. I) Circulating water bath. J) Cannulation needle with heart, attached to the perfusion outlet port. Please click here to view a larger version of this figure.
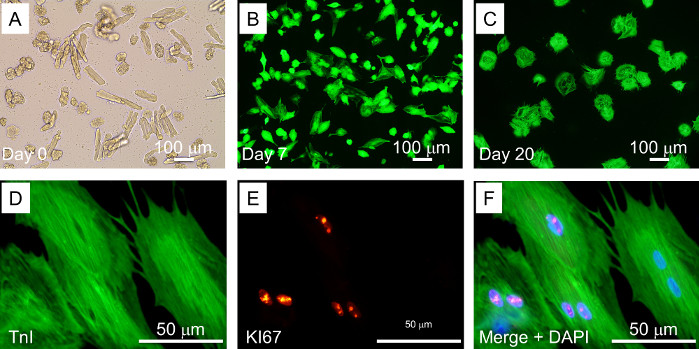
Figure 2: Adult rat CM isolation, transfection, and long-term culture. A) Adult rat CM, immediately after isolation. B) Adult rat CM on Day 7 after isolation. C) Adult rat CM on day 20. D-F) KI67 positive rat CM on day 7 after siRb1+siMeis2 transfection. Troponin-I = green; DAPI = blue; KI67 = red. All the experiments were performed in duplicate and repeated at least three times. Please click here to view a larger version of this figure.
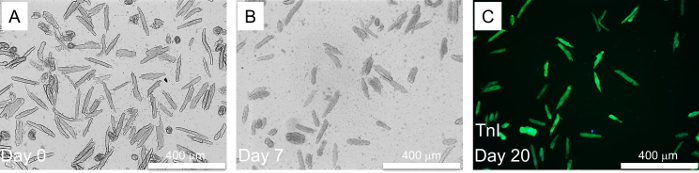
Figure 3: Adult mouse CM isolation and long term culture. A) Adult mice CM, immediately after isolation. B) Adult mice CM on Day 7 after isolation. C) Adult mice CM stained with Cardiac Troponin-I = green on day 20. Please click here to view a larger version of this figure.
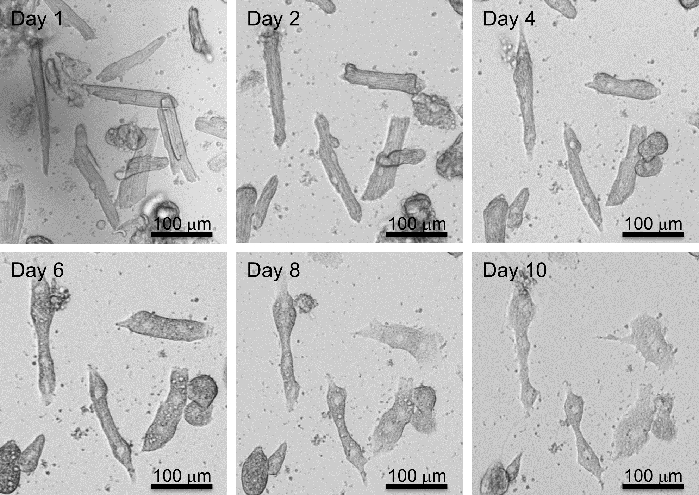
Figure 4: Adult mouse CM de-differentiation. Adult mice CM showing morphological changes during long-term culture (day 0 to Day 10) in DMEM-HG media, supplemented with 10%FBS. Please click here to view a larger version of this figure.
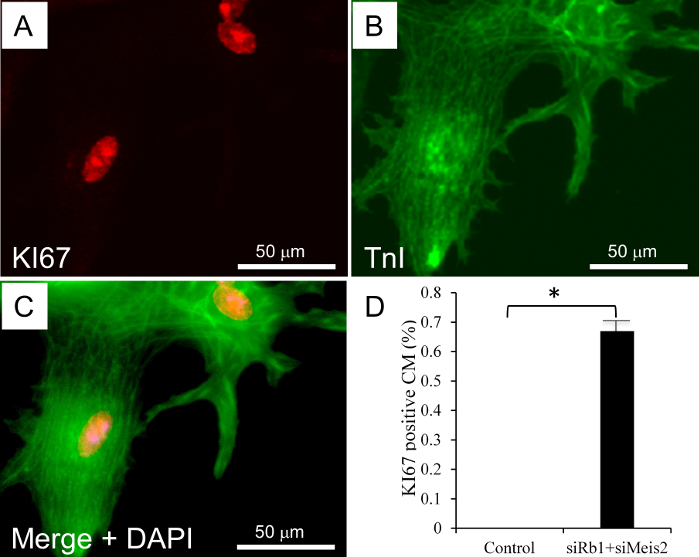
Figure 5: Adult mouse CM transfection and proliferation. A-C) KI67 positive mouse CM on day 10 after siRb1+siMeis2 transfection. Troponin-I = green; DAPI = blue; KI67 = red. D) Bar graph shows a significant increase in KI67 positive adult mouse CMs in the siRNA-cocktail transfected group versus control. Results are presented as mean±SEM; * = p-value ≤0.05. p-value ≤0.05 was considered statistically significant. All the experiments were performed in triplicate (n=3 Male,3 Female). Please click here to view a larger version of this figure.
