Зондирующая динамика мягкого вещества с нейтронами
Исследование динамических свойств белков и пептидов является важной частью биофизических исследований, и сегодня существует множество хорошо разработанных методов для доступа к широкому спектру энергетических ландшафтов1. Соотнесение экспериментально выявленной динамики белков с их биологической функцией является гораздо более сложной задачей, требующей сложных математических моделей и компьютерного моделирования динамики. Важность нейтронной спектроскопии для анализа движений белка была подчеркнута в нескольких хорошо принятых и широко признанных исследованиях 1,2,3,4,5. Прежде чем исследовать разнообразный энергетический ландшафт внутренней динамики белка, требуется краткий обзор динамических процессов в мягкой материи и того, как нейтроны могут получить к ним доступ.
Чувствительность нейтронов к изотопной конфигурации и тип взаимодействий, которые они проявляют с мягким веществом, делает рассеяние нейтронов одним из наиболее универсальных методов исследования6. Существует широкий спектр шкал длин корреляции и времени корреляции, к которым могут получить доступ нейтроны, от ядерных возбуждений и атомных колебаний до коллективных движений и медленных процессов релаксации, таких как изотропные вращения и диффузионные движения. При исследовании рассеянных нейтронов на предмет их переноса энергии можно выделить три основных взаимодействия: упругое рассеяние, при котором отсутствует энергообмен между входящим нейтроном и частицей в образце; неупругое рассеяние с большим, поддающимся количественной оценке энергетическим обменом между нейтроном и частицей; и своеобразный случай квазиупругого рассеяния, обозначающий очень малый перенос энергии по сравнению с падающей энергией нейтронов 1,7. Эти взаимодействия дают точную информацию об исследуемом материале и составляют теоретическую основу широкого спектра методов рассеяния нейтронов.
При упругом рассеянии детектор записывает направления нейтронов как дифракционную картину, которая показывает положение атомов образца относительно друг друга. Получена информация о корреляциях атомных положений (т.е. интегральная интенсивность S(Q) относительно переноса импульса Q, относящаяся только к структурной информации). Этот принцип лежит в основе дифракции нейтронов8.
Сложность возникает, когда передача энергии больше не равна нулю из-за возбуждений и внутренних флуктуаций в материале образца. Это составляет основу нейтронной спектроскопии, в которой рассеянные нейтроны исследуются как функция как переноса энергии E , так и переноса импульса Q. Получена динамическая и структурная информация. Нейтронная спектроскопия измеряет одинаковую интегральную интенсивность S(Q) для передачи энергии (т.е. изменение скорости нейтронов из-за рассеяния образцов, S(Q,ω) = S(Q, E), что также называется коэффициентом динамической структуры)9.
Для расчета рассеяния из материала более адекватно использовать парную корреляционную функцию 7,10. В дифракционном случае статическая парная корреляционная функция G(r) дает вероятность нахождения центра частицы на заданном расстоянии r от центра другой частицы. Спектроскопия обобщает статическую парную корреляционную функцию и включает энергию/частоту/время в уравнение рассеяния. Парная корреляционная функция G(r) становится функцией времени G(r, t), которая может быть разложена на отдельную корреляционную функцию пары атомов GD(r, t) и самокорреляционную функцию GS(r, t). Они описывают два типа корреляций: парно-коррелированные движения атомов, которые управляют когерентным рассеянием, и самокорреляция, которая управляет некогерентным рассеянием10.
Когерентное рассеяние является рассеянием от «среднего» и зависит от относительной фазы рассеянных волн. В малоугловом режиме рассеяния рассеянные нейтронные волны от разных центров рассеяния (разных атомов) конструктивно взаимодействуют (имеют сходные фазы), а коллективное движение атомов наблюдается с сильным усилением интенсивности. Когерентное рассеяние по существу описывает рассеяние одного нейтрона от всех ядер в образце10.
Когда между рассеянными нейтронными волнами из разных центров не возникает конструктивной интерференции, во времени следует один атом, и наблюдается самокорреляция между положением атома в момент времени t = 0 и того же атома в момент времени t. Таким образом, информация об относительных положениях атомов теряется, а фокус делается только на локальных флуктуациях. Рассеяние от локальных флуктуаций управляет некогерентным рассеянием. Некогерентное рассеяние изотропно, способствует фоновому сигналу и ухудшает сигнал-шум10,11.
Объединив все вышеперечисленное, мы выделяем четыре основных процесса рассеяния нейтронов10: (1) упругий когерентный (измеряет корреляции атомных положений), (2) неупругий когерентный (измеряет коллективные движения атомов), (3) упругий некогерентный (способствует фону, уменьшает интенсивность рассеяния на коэффициент Дебая-Уоллера (DWF) и измеряет коэффициент упругой некогерентной структуры (EISF), описывая геометрию диффузионных движений в замкнутой геометрии, и (4) неупругий некогерентный (измеряет динамику одного атома и самокорреляцию).
Динамические процессы, к которым нейтроны могут получить доступ в биологии, варьируются от демпфирования низкочастотных атомных и молекулярных колебаний, взаимодействия молекул растворителей с биоповерхностями и диффузионных процессов в гидратационном слое макромолекул и ограниченной геометрии до прямых поступательных, вращательных и кувыркающихся диффузных движений, белковых доменов и аллостерических движений1 . Широкое разнообразие нейтронных методов и приборов для измерения динамики белка основано на том, как достигается ахроматизация падающего или исходящего пучка нейтронов и как выполняется энергетический анализ рассеянных нейтронов. От трехосевых до спектрометров времени полета, обратного рассеяния и спин-эхо можно исследовать динамические процессы с характерным временем от 1 х 10-14 с до 1 х 10-6 с (фемтосекунды до микросекунд)12.
Национальная лаборатория Оук-Ридж с двумя известными источниками нейтронов, источником нейтронов Spallation – SNS13 и реактором с высоким изотопным потоком – HFIR14, имеет один из лучших наборов спектрометров для исследования динамики в биоматериалах. Некоторые из наиболее красноречивых примеров включают использование спектрометра холодного нейтронного измельчителя (CNCS) в SNS15 для исследования динамического возмущения гидратационной воды вокруг зеленого флуоресцентного белка в растворе16 или субпикосекундных коллективных колебаний нескольких белков17. Повторяющаяся проблема исследований неупругого рассеяния нейтронов заключается в том, что некоторые биологические процессы слишком медленны, чтобы их можно было наблюдать. Без экстремальных установок, которые приводят к огромной потере интенсивности нейтронов, спектрометры времени пролета ограничены разрешением энергии 10 мкЭВ, что соответствует максимальной шкале времени ~ 200 пс10,11. Этого недостаточно для наблюдения крупномасштабных движений в белках. Поэтому часто требуются приборы с более высоким энергетическим разрешением, такие как спектрометры обратного рассеяния. Сочетание методов времени полета и обратного рассеяния оказалось мощным для исследования изменения внутренней динамики цитохрома P450cam (CYP101), фермента, который катализирует гидроксилирование камфоры18.
Микроскопическая диффузия, измеренная спектрометром обратного рассеяния в SNS-BASIS19 , была удивительно хорошо определена и могла быть разделена на диффузионность воды (гидратационная, цитоплазматическая и объемная вода) и диффузионность клеточных компонентов у плоских плоских червей, первого живого животного, изучаемого рассеянием нейтронов20 . Обратное рассеяние является спектроскопическим методом с высоким разрешением, но оно также ограничено несколькими мкэВ = несколькими наносекундами, в то время как медленная динамика в биоматериалах также проявляется как время выживания корреляции между положением атома или ориентацией спина (например, релаксационные процессы, которые регулярно происходят в диапазоне времени от десяти до сотен наносекунд).
Нейтронная спиновая эхо-спектроскопия (NSE) является единственным методом рассеяния нейтронов, достигающим такого высокого разрешения. В отличие от других нейтронных методов, NSE не требует ахроматизации пучка, поскольку использует квантово-механическую фазу нейтронов, которая является их магнитными моментами. Манипулирование магнитными моментами позволяет использовать широкое распределение длин волн пучка нейтронов, при этом методика чувствительна к очень малым изменениям скорости нейтронов порядка 1 х 10-4. NSE был успешно использован для исследования медленной динамики белков в растворе для многих белков. Среди этих многочисленных новаторских исследований мы признаем изучение сегментарной гибкости иммуноглобулина свиней21; связанные доменные движения в Taq полимеразе22; доменные движения в тетрамере дрожжевого спиртдегидрогеназы23; изменение конформации в фосфоглицераткиназе при связывании субстрата3; активация доменных движений и динамическое распространение аллостерических сигналов в белке регуляторного кофактора обмена Na+/H+ 1 (NHERF1) 4,24,25; динамика компактного состояния ртутной ионредуктазы26; и диффузия гемоглобина в эритроцитах27. Два более поздних исследования в области динамики белка выявили гибкость антитела человека Иммуноглобулина G (IgG) в качестве энтропийного источника28 и характеристики вклада растворителя в динамику внутренне неупорядоченного основного белка миелина (MBP)5.
В настоящей статье объясняются основные принципы NSE, множественные подготовительные методы, рекомендуемые для тщательного исследования динамики белка, а также методология и экспериментальный протокол сбора данных NSE на спектрометре NSE в SNS, SNS-NSE. Протокол характеризует два белка: IgG, обычный белок антител человека, и внутренне неупорядоченный белок MBP. Биофизические последствия, исследовательская актуальность примеров и ограничения метода обсуждаются кратко.
NSE спектроскопия, метод измерения медленной динамики
NSE – это поляризованный метод, который использует нейтронное время пролета для измерения обмена энергией (потери поляризации) из-за квазиупругого взаимодействия между нейтронами и атомами в образце. В основе спектроскопии NSE лежат два основных принципа: (1) способность спина нейтрона прецессировать в магнитном поле с частотой, пропорциональной магнитной силе, а именно частоте 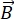 Лармора29, и (б) спин-эхо или эхо Ханна, представляющее собой манипулирование и перефокусировку поляризационного сигнала при подаче серии радиочастотных импульсов30.
Лармора29, и (б) спин-эхо или эхо Ханна, представляющее собой манипулирование и перефокусировку поляризационного сигнала при подаче серии радиочастотных импульсов30.
Основы процесса NSE можно резюмировать в нескольких простых шагах 6,11, используя рисунок 1. (1) Пучок нейтронов, создаваемый источником (положение 1), поляризован (положение 2), направляется и транспортируется (положение 3) и поступает на вход в спектрометр NSE, где он поворачивается на 90° первым пи-полуфлаппером (положение 4). (2) Поляризованный пучок (например, магнитные моменты нейтронов) становится перпендикулярным линиям магнитного поля первого магнита (первая зона прецессии, положение 5) и начинает прецессировать. (3) На конце магнита спины нейтронов накапливают определенный угол прецессии, пропорциональный напряженности магнитного поля и времени полета, проведенному внутри (в основном обратно пропорционален скорости нейтрона). Отдельные скорости нейтронов кодируются в пределах угла прецессии в конце первой зоны прецессии. (4) Вблизи положения образца пи-флиппер (положение 6) меняет ориентацию спина на 180°, изменяя знак угла прецессии. (5) Нейтроны взаимодействуют с молекулами образца (позиция 7) и рассеиваются. (6) Рассеянные нейтроны входят и прецессируют во второй зоне прецессии (позиция 8), но становятся обратно ориентированными. (7) Другой пи-полуплащ (положение 9) используется для поворота ориентации спина от перпендикулярного к горизонтальному направлению. Это остановит прецессию, переводя угол прецессии φ в поляризацию, пропорциональную cos(φ). (8) Анализатор (позиция 10) выбирает нейтроны на основе одной ориентации. Если взаимодействие с образцом упругое, скорость нейтрона не изменится. Нейтроны будут проводить одинаковое количество времени, пролетая в первой и второй зонах прецессии, а накопленные углы прецессии полностью восстанавливаются. Полная поляризация восстанавливается на детекторе (позиция 11) как эхо исходной поляризации (т.е. спин-эхо). (9) Однако в NSE рассеяние является квазиупругим, поэтому небольшой обмен энергией между нейтронами и молекулами образца приводит к различным скоростям нейтронов после рассеяния образцом. Из-за различных скоростей нейтроны проведут дополнительное время, пролетая через вторую зону прецессии, и не смогут должным образом восстановить свой угол прецессии. На детекторе извлекается частичная поляризация, а потеря поляризации из-за спиновой релаксации пропорциональна кос-фурье-преобразованию спектральной функции S(Q, ω), промежуточной функции рассеяния F(Q, t). (10) Параметр времени функции F(Q, t) пропорционален напряженности магнитного поля прецессии. Сканирование потери поляризации в зависимости от напряженности магнитного поля дает, следовательно, релаксационную функцию, которая зависит от динамических процессов внутри образца.

Рисунок 1: Фотография спектрометра NSE в SNS (SNS-NSE) и схемы траектории полета нейтронов с наиболее важными функциональными компонентами. Справа налево: 1 = источник нейтронов; 2 = система измельчителей-бендеров-поляризаторов-вторичных затворов; 3 = направляющие для транспортировки луча; 4 = пи/2 флиппера для первого вращения на 90°; 5 = первая зона прецессии; 6 = пи-флиппер для вращения на 180°; 7 = площадь образца и среда отбора проб (здесь показана криопечь); 8 = вторая зона прецессии; 9 = пи/2 флиппера для второго вращения на 90°; 10 = анализатор; 11 = детектор. (Обратите внимание, что части 3, а также 2 и 1 расположены за синей стеной внутри экранирования; измельчители заменены селектором скорости для NSE на основе реактора). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
