Dinámica de sondeo de la materia blanda con neutrones
La investigación de las propiedades dinámicas de las proteínas y los péptidos es una parte importante de la investigación biofísica, y hoy en día existen muchos métodos bien desarrollados para acceder a una amplia gama de paisajes energéticos1. Relacionar la dinámica revelada experimentalmente de las proteínas con su función biológica es una tarea mucho más difícil, que requiere modelos matemáticos complejos y simulaciones dinámicas asistidas por computadora. La importancia de la espectroscopia de neutrones para el análisis de los movimientos de las proteínas ha sido enfatizada en varios estudios bien recibidos y ampliamente reconocidos 1,2,3,4,5. Antes de explorar el diverso panorama energético de la dinámica interna de proteínas, se requiere una breve descripción de los procesos dinámicos en la materia blanda y cómo los neutrones pueden acceder a ellos.
La sensibilidad de los neutrones a la configuración isotópica y el tipo de interacciones que muestran con la materia blanda hace que la dispersión de neutrones sea una de las técnicas de investigación más versátiles6. Existe un amplio espectro de escalas de longitud de correlación y tiempos de correlación a los que los neutrones pueden acceder, desde excitaciones nucleares y vibraciones atómicas hasta movimientos colectivos y procesos de relajación lentos como rotaciones isotrópicas y movimientos difusivos. Al investigar los neutrones dispersos para su transferencia de energía, se pueden distinguir tres interacciones principales: la dispersión elástica, en la que no hay intercambio de energía entre el neutrón entrante y la partícula en la muestra; la dispersión inelástica, con un gran intercambio de energía cuantificable entre neutrones y partículas; y el peculiar caso de la dispersión cuasielástica que designa una transferencia de energía muy pequeña en comparación con la energía neutrónica incidente 1,7. Estas interacciones proporcionan información precisa sobre el material investigado y forman la base teórica de una amplia variedad de técnicas de dispersión de neutrones.
En la dispersión elástica, el detector registra las direcciones de los neutrones como un patrón de difracción, que muestra la posición de los átomos de muestra entre sí. Se adquiere información sobre las correlaciones de las posiciones atómicas (es decir, la intensidad integrada S (Q) con respecto a la transferencia de momento Q, que se refiere solo a la información estructural). Este principio constituye la base de la difracción de neutrones8.
La complejidad surge cuando la transferencia de energía ya no es cero debido a excitaciones y fluctuaciones internas en el material de la muestra. Esto forma la base de la espectroscopia de neutrones, en la que los neutrones dispersos se investigan en función tanto de la transferencia de energía E como de la transferencia de momento Q. Se obtiene información dinámica y estructural. La espectroscopia de neutrones mide la misma intensidad integrada S(Q) para la transferencia de energía (es decir, el cambio de velocidad de los neutrones debido a la dispersión de la muestra, S(Q,ω) = S(Q, E), que también se conoce como el factor de estructura dinámica)9.
Para calcular la dispersión a partir de un material, es más adecuado utilizar la función de correlación de pares 7,10. En el caso de difracción, la función de correlación de pares estáticos G(r) da la probabilidad de encontrar el centro de una partícula a una distancia dada r del centro de otra partícula. La espectroscopia generaliza la función de correlación de pares estáticos e incluye energía/frecuencia/tiempo en la ecuación de dispersión. La función de correlación de pares G(r) se convierte en una función del tiempo G(r, t), que puede descomponerse en una función de correlación de pares de átomos distinta GD(r, t), y una función de autocorrelación GS(r, t). Estos describen dos tipos de correlaciones: movimientos correlacionados por pares de átomos que gobiernan la dispersión coherente y autocorrelación que gobierna la dispersión incoherente10.
La dispersión coherente es la dispersión de “la media” y depende de la fase relativa de las ondas dispersas. En el régimen de dispersión de ángulo pequeño, las ondas de neutrones dispersas de diferentes centros de dispersión (diferentes átomos) interfieren constructivamente (tienen fases similares), y el movimiento colectivo de los átomos se observa con una fuerte mejora de intensidad. La dispersión coherente describe esencialmente la dispersión de un solo neutrón de todos los núcleos de la muestra10.
Cuando no se produce ninguna interferencia constructiva entre las ondas de neutrones dispersas de diferentes centros, se sigue un solo átomo en el tiempo, y se observa la autocorrelación entre la posición del átomo en el tiempo t = 0 y el mismo átomo en el tiempo t. Por lo tanto, la información sobre las posiciones relativas de los átomos se pierde, y el enfoque está solo en las fluctuaciones locales. La dispersión de las fluctuaciones locales gobierna la dispersión incoherente. La dispersión incoherente es isotrópica, contribuye a la señal de fondo y degrada la señal a ruido10,11.
Combinando todo lo anterior, distinguimos cuatro procesos principales de dispersión de neutrones10: (1) coherente elástico (mide las correlaciones de las posiciones atómicas), (2) coherente inelástico (mide los movimientos colectivos de los átomos), (3) elástico incoherente (contribuye al fondo, reduce la intensidad de dispersión por el factor de Debye-Waller (DWF) y mide el factor de estructura incoherente elástica (EISF), describiendo la geometría de los movimientos difusivos en geometría confinada, y (4) incoherente inelástica (mide la dinámica de un solo átomo y la autocorrelación).
Los procesos dinámicos a los que los neutrones pueden acceder en biología van desde la amortiguación de vibraciones atómicas y moleculares de baja frecuencia, la interacción de moléculas de solvente con biosuperficies y procesos de difusión en la capa de hidratación de macromoléculas y geometría confinada, hasta movimientos difusivos de traslación, rotación y caída de corto alcance, y dominios de proteínas y movimientos alostéricos1 . La amplia diversidad de métodos e instrumentos de neutrones para medir la dinámica de las proteínas se basa en cómo se logra la acromatización del haz de neutrones incidente o saliente y cómo se realiza el análisis energético de los neutrones dispersos. Desde el triple eje hasta el tiempo de vuelo, la retrodispersión y los espectrómetros de espín-eco, se pueden explorar procesos dinámicos con tiempos característicos entre 1 x 10-14 s y 1 x 10-6 s (femtosegundos a microsegundos)12.
El Laboratorio Nacional de Oak Ridge, con sus dos fuentes de neutrones de renombre, la Fuente de Neutrones de Espalación – SNS13 y el Reactor de Alto Flujo de Isótopos – HFIR14, tiene uno de los mejores conjuntos de espectrómetros para investigar la dinámica en biomateriales. Algunos de los ejemplos más elocuentes incluyen el uso del espectrómetro de picador de neutrones fríos (CNCS) en SNS15 para investigar la perturbación dinámica del agua de hidratación alrededor de la proteína fluorescente verde en la solución16 o las vibraciones colectivas sub-picosegundos de varias proteínas17. Un problema recurrente de las investigaciones de dispersión de neutrones inelásticos es que algunos procesos biológicos son demasiado lentos para ser observados. Sin configuraciones extremas que conduzcan a una gran pérdida de intensidad de neutrones, los espectrómetros de tiempo de vuelo están limitados a una resolución de energía de 10 μeV, lo que corresponde a una escala de tiempo máxima de ~ 200 ps10,11. Esto no es suficiente para observar movimientos a gran escala en las proteínas. Por lo tanto, a menudo se necesitan instrumentos con una mayor resolución de energía, como los espectrómetros de retrodispersión. La combinación de las técnicas de tiempo de vuelo y retrodispersión ha demostrado ser poderosa para investigar el cambio en la dinámica interna del citocromo P450cam (CYP101), una enzima que cataliza la hidroxilación alcanfor18.
La difusividad microscópica medida por el espectrómetro de retrodispersión en SNS-BASIS19 estaba sorprendentemente bien definida y podía separarse en la difusividad del agua (hidratación, citoplasmática y agua a granel) y la difusividad de los constituyentes celulares en gusanos planos planarios, el primer animal vivo que se estudió mediante dispersión de neutrones20 . La retrodispersión es una técnica espectroscópica de alta resolución, pero también se limita a varios μeV = varios nanosegundos, mientras que la dinámica lenta en los biomateriales también se manifiesta como el tiempo de supervivencia de la correlación entre la posición atómica o las orientaciones de espín (por ejemplo, procesos de relajación, que ocurren regularmente en el rango de tiempo de diez a cientos de nanosegundos).
La espectroscopia de eco de espín de neutrones (NSE) es la única técnica de dispersión de neutrones que alcanza una resolución tan alta. A diferencia de otras técnicas de neutrones, NSE no requiere la acromatización del haz, ya que utiliza la fase mecánica cuántica de los neutrones, que son sus momentos magnéticos. La manipulación de momentos magnéticos permite el uso de una amplia distribución de longitud de onda del haz de neutrones, mientras que la técnica es sensible a cambios de velocidad de neutrones muy pequeños del orden de 1 x 10-4. NSE se ha utilizado con éxito para investigar la dinámica lenta de las proteínas en solución para muchas proteínas. Entre estos muchos estudios pioneros, reconocemos el estudio de la flexibilidad segmentaria de la inmunoglobulina porcina21; los movimientos de dominio acoplado en la polimerasaTaq 22; los movimientos del dominio en el tetrámero de levadura alcohol deshidrogenasa23; el cambio de conformación en la fosfoglicerato quinasa sobre la unión al sustrato3; la activación de los movimientos de dominio y la propagación dinámica de señales alostéricas en la proteína 4,24,25 del cofactor regulador del intercambio Na+/H+ (NHERF1); la dinámica de un estado compacto de iones mercúricos reductasa26; y la difusión de hemoglobina en los glóbulos rojos27. Dos estudios más recientes en dinámica de proteínas han expuesto la flexibilidad del anticuerpo humano Inmunoglobulina G (IgG) como primavera entrópica28 y las características de la contribución de disolventes a la dinámica de la proteína básica de mielina intrínsecamente desordenada (MBP)5.
El presente artículo explica los principios básicos de NSE, los múltiples métodos preparatorios recomendados para una investigación exhaustiva de la dinámica de proteínas, así como la metodología y el protocolo experimental para la adquisición de datos de NSE en el espectrómetro NSE en SNS, SNS-NSE. El protocolo caracteriza dos proteínas: IgG, una proteína de anticuerpos humanos regulares, y la proteína intrínsecamente desordenada MBP. Las implicaciones biofísicas, la relevancia de la investigación de los ejemplos y las limitaciones de la técnica se discuten brevemente.
Espectroscopia NSE, el método para mediciones de dinámica lenta
NSE es una técnica polarizada que utiliza el tiempo de vuelo de neutrones para medir el intercambio de energía (pérdida de polarización) debido a la interacción cuasi-elástica entre neutrones y átomos en una muestra. En el núcleo de la espectroscopia NSE se encuentran dos principios básicos: (1) la capacidad del espín de neutrones para precesar en el campo magnético con una frecuencia proporcional a la fuerza 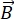 magnética, a saber, la frecuencia de Larmor29, y (b) el espín-eco o eco de Hann, que representa la manipulación y reenfoque de la señal de polarización al aplicar una serie de pulsos de radiofrecuencia30.
magnética, a saber, la frecuencia de Larmor29, y (b) el espín-eco o eco de Hann, que representa la manipulación y reenfoque de la señal de polarización al aplicar una serie de pulsos de radiofrecuencia30.
Los conceptos básicos del proceso NSE se pueden resumir en unos sencillos pasos 6,11 utilizando la Figura 1. (1) El haz de neutrones producido por la fuente (posición 1) es polarizado (posición 2), guiado y transportado (posición 3), y llega a la entrada del espectrómetro NSE, donde se gira 90° por la primera aleta pi-half (posición 4). (2) El haz polarizado (por ejemplo, momentos magnéticos de neutrones) se vuelve perpendicular a las líneas del campo magnético del primer imán (primera zona de precesión, posición 5) y comienza a precesar. (3) Al final del imán, los espines de neutrones acumulan un cierto ángulo de precesión proporcional a la intensidad del campo magnético y al tiempo de vuelo pasado en su interior (básicamente inversamente proporcional a la velocidad del neutrón). Las velocidades de neutrones individuales están codificadas dentro de su ángulo de precesión al final de la primera zona de precesión. (4) Cerca de la posición de la muestra, el pi-flipper (posición 6) invierte la orientación del espín en 180°, cambiando el signo del ángulo de precesión. (5) Los neutrones interactúan con las moléculas de la muestra (posición 7) y se dispersan. (6) Los neutrones dispersos entran y precesan en la segunda zona de precesión (posición 8) pero se orientan hacia atrás. (7) Se utiliza otra aleta pi-half (posición 9) para girar la orientación del giro de perpendicular a la dirección horizontal. Esto detendrá la precesión, traduciendo el ángulo de precesión φ en polarización proporcional a cos(φ). (8) El analizador (posición 10) selecciona los neutrones en función de una orientación. Si la interacción con la muestra es elástica, la velocidad del neutrón no cambiará. Los neutrones pasarán una cantidad idéntica de tiempo volando en la primera y segunda zonas de precesión, y los ángulos de precesión acumulados se recuperan por completo. La polarización completa se restaura en el detector (posición 11) como un eco de la polarización original (es decir, espín-eco). (9) Sin embargo, en NSE, la dispersión es cuasi elástica, por lo que un pequeño intercambio de energía entre neutrones y moléculas de muestra conduce a diferentes velocidades de neutrones después de la dispersión por la muestra. Debido a las diferentes velocidades, los neutrones pasarán un tiempo adicional volando a través de la segunda zona de precesión y no habrán recuperado adecuadamente su ángulo de precesión. Se recupera una polarización parcial en el detector, y la pérdida de polarización debido a la relajación del espín es proporcional a la transformada cos-Fourier de la función espectral S(Q, ω), la función de dispersión intermedia F(Q, t). (10) El parámetro de tiempo de la función F(Q, t) es proporcional a la intensidad del campo magnético de precesión. El escaneo de la pérdida de polarización en función de la intensidad del campo magnético produce, por lo tanto, una función de relajación que depende de los procesos dinámicos dentro de la muestra.

Figura 1: Fotografía del espectrómetro NSE en SNS (SNS-NSE) y esquema de trayectoria de vuelo de neutrones con los componentes funcionales más importantes. De derecha a izquierda: 1 = fuente de neutrones; 2 = choppers-bender-polarizador-sistema de obturador secundario; 3 = guías de transporte de vigas; 4 = pi/2 flipper para el primer giro de 90°; 5 = primera zona de precesión; 6 = pi flipper para 180° spin-turn; 7 = área de muestra y entorno de muestra (aquí, se muestra el crio-horno); 8 = segunda zona de precesión; 9 = pi/2 flipper para el segundo giro de 90°; 10 = analizador; 11 = detector. (Tenga en cuenta que las porciones de 3, así como 2 y 1, están situadas detrás de la pared azul dentro del blindaje; los helicópteros se reemplazan por un selector de velocidad para NSE basado en reactores). Haga clic aquí para ver una versión más grande de esta figura.
