Metamerisk segmentering af organismer er meget udbredt i naturen. Gentagne strukturer er afgørende for funktionaliteten af laterale organer som hvirvler, muskler, nerver, fartøjer, lemmer eller blade i en kropsplan1. Som et resultat af sådanne fysiologiske og geometriske begrænsninger af den aksiale symmetri udviser de fleste karinaer af Bilateria- såsom annelider, leddyr og akkordater-udstilling segmentering af deres embryonale væv (f.eks. ectoderm, mesoderm) antero-posteriorly.
Hvirveldyr embryoner sekventielt segment deres paraxial mesoderm langs de store organ akse i somites med artsspecifikke intervaller, tæller, og størrelse distributioner. På trods af en sådan robusthed blandt individuelle embryoner inden for en art er somitsegmentering alsidig mellem hvirveldyrarter. Segmentering sker i et stort regime af tidsintervaller (fra 25 min i zebrafisk til 5 timer hos mennesker), størrelser (fra ~ 20 μm i hale somites af zebrafisk til ~ 200 μm i trunk somites af mus) og tæller (fra 32 i zebrafisk til ~ 300 i majsslanger)2. Mere interessant er det, at fiskefostre kan udvikle sig i en lang række temperaturer (fra ~20,5 °C op til 34 °C for zebrafisk), samtidig med at deres somitter bevares med korrekt størrelsesfordeling ved at kompensere for både segmenteringsintervaller og aksiale forlængelseshastigheder. Ud over sådanne interessante træk forbliver zebrafisk som en nyttig modelorganisme til at studere segmentering i hvirveldyr på grund af den eksterne, synkrone og gennemsigtige udvikling af en mangfoldighed af søskendefostre samt deres tilgængelige genetiske værktøjer. Fra mikroskopisk perspektiv udvikler teleostembryoner sig på en voluminøs sfærisk æggeblomme, der strækker sig og afrunder det gastrulerende væv omkring det(figur 1A). I denne artikel præsenterer vi en flad 3-D vævsudgravningskultur til zebrafiskhaler. Dette strålesystem omgår æggeblommemassens sfæriske begrænsninger, hvilket giver adgang til levende billeddannelse af fiskeembryoner i høj opløsning med henblik på somite-mønstre.
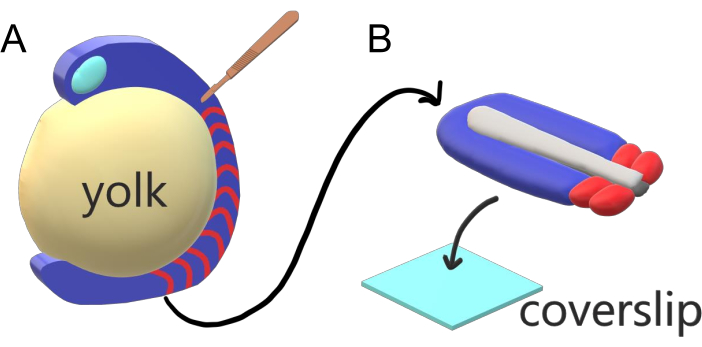
Figur 1: Slide Chamber Explant System for Zebrafish Embryos. (A) Zebrafiskembryoner har fordele for levende billeddannelse, såsom gennemsigtigheden af gastrulerende embryonalt væv (blåt), men vævet dannes omkring en voluminøs sfærisk æggeblommemasse (gul), som forhindrer næsten objektiv billeddannelse i høj opløsning i intakte embryoner. Hale explants kan dissekeres begyndende med en mikrokirurgisk kniv (brun) skåret fra vævet foran somites (rød) og fortsætter ved grænsen til æggeblommen posteriorly. (B) Dissekerede hale explants kan placeres på en coverslip (lyseblå) dorsoventrally; holde neuralt væv (lysegrå) på toppen og notochord (mørkegrå) i bunden. Klik her for at se en større version af dette tal.
