Метамерная сегментация организмов широко используется в природе. Повторяющиеся структуры необходимы для функциональности боковых органов, таких как позвонки, мышцы, нервы, сосуды, конечности или листья в плане тела1. В результате таких физиологических и геометрических ограничений осевой симметрии большинство типов Bilateria, таких как кольчатые черви, членистоногие и хордовые, демонстрируют сегментацию своих эмбриональных тканей (например, эктодермы, мезодермы) с задней стороны.
Эмбрионы позвоночных последовательно сегментируют свою параксиальную мезодерму вдоль главной оси тела в сомиты с видоспецифичными интервалами, подсчетами и распределением размеров. Несмотря на такую устойчивость среди отдельных эмбрионов внутри вида, сегментация сомита универсальна между видами позвоночных. Сегментация происходит в обширном режиме временных интервалов (от 25 мин у рыбок данио до 5 ч у человека), размеров (от ~20 мкм в хвостовых сомитах рыбок данио до ~200 мкм у хоботных сомитов мышей) и подсчетов (от 32 у рыбок данио до ~300 у кукурузных змей)2. Что еще более интересно, эмбрионы рыб могут развиваться в широком диапазоне температур (от ~ 20,5 ° C до 34 ° C для рыбок данио), сохраняя при этом свои сомиты нетронутыми с правильным распределением размеров, компенсируя как интервалы сегментации, так и скорости осевого удлинения. Помимо таких интересных особенностей, рыбки данио остаются полезным модельным организмом для изучения сегментации у позвоночных из-за внешнего, синхронного и прозрачного развития обильной популяции эмбрионов братьев и сестер, а также их доступных генетических инструментов. Неблагоприятно с точки зрения микроскопии, телеостные эмбрионы развиваются на громоздком сферическом желтке, растягивая и округляя гаструлирующие ткани вокруг него(рисунок 1А). В этой статье мы представляем сплющенный 3-D тканевый эксплант культуры для хвостов рыбок данио. Эта эксплантная система обходит сферические ограничения желточной массы, позволяя получить доступ к живым изображениям эмбрионов рыб с высоким разрешением для сомит-паттерна.
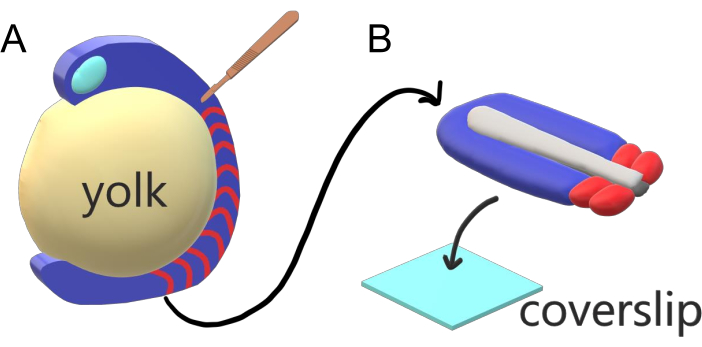
Рисунок 1:Slide Chamber Explant System for Zebrafish Embryos. (A) Эмбрионы рыбок данио имеют преимущества для живой визуализации, такие как прозрачность гаструлирующей эмбриональной ткани (синий), но ткань образуется вокруг громоздкой сферической желточной массы (желтый), которая предотвращает почти объективную визуализацию с высоким разрешением у неповрежденных эмбрионов. Хвост эксплантов можно рассечь, начиная с микрохирургического ножа (коричневого), вырезанного из ткани с передней части сомитов (красного) и продолжая на границе с желтком с задней стороны. (B) Рассеченные хвостовые экспланты могут быть размещены на покровном (светло-синем) дорсовентральном; сохранение нервной ткани (светло-серого) сверху и нотохорды (темно-серого) внизу. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
