二部GAL4-UASシステムは、昆虫モデルの生物ショウジョウバエメラノガスター1,2,3における遺伝子の機能的特徴付けの役馬である。GAL4-UASシステムを使用するには、トランスジェニックドライバラインは、調節配列の制御下にある酵母転写因子GAL4を発現し、GAL4によって認識される上流活性化シーケンス(UAS)によって制御される目的の遺伝子またはRNA干渉(RNAi)構築物を運ぶ応答線と交差する。この十字架の子孫は、GAL4発現を制御するプロモーターによって指示される時空間パターンにおける関心のトランスジーンを表す(図1)。ドライバー応答者十字の子孫によって表示される型のフェノタイプは、候補遺伝子の機能を解明するために評価することができる。D.メラノガスターは、他の生物から遺伝子を調べるために使用されているが4,5,6,7,, GAL4-UASシステムは、関心のある種8,9,11,12,13,14の直接分析を提供するために、医療および農業上重要の昆虫での使用に適応されています。
アフリカのマラリア蚊、アノフェレスガンビアでは、GAL4-UASシステムが最初に細胞株共トランスフェクション9によってテストされました。複数の構成体が異なる対の組み合わせで効率のためにアッセイされ、14のタンデム反復UASが小型の人工イントロン(UAS-14i)を補い、GAL4ドライバのパネルで使用すると、最も広い範囲の活性化電位を示すことがわかった。インビボ機能を実証するために、これらの構成体は、ピギーバクトトランスフォーメーション8によって2つの別々のトランスジェニックAn.ガンビアラインを作成するために使用されました:midgut特異的プロモーターによって駆動されるGAL4を運ぶドライバーラインと、UAS配列の調節下にあるルシメラーゼおよび強化された黄色蛍光タンパク質(eYFP)遺伝子の両方を含む応答線。腸特異的ルシメラーゼ活性および生前性における蛍光は、このシステムがアノフェレスで効率的であることを示した。それ以来、オエノサイト15やヘモサイト16を含むベクター容量および殺虫剤耐性に重要な他の組織におけるトランス遺伝子を発現するドライバラインが作成され、かつ、ユビキタスパターン10に近いものである。また、代謝および隔離媒介性殺虫剤耐性に関与すると考えられるアッセイ遺伝子、カチカリン炭化水素合成、および異なる細胞および組織タイプに蛍光タグを付ける測定遺伝子にも多数のUASラインが生成されている(表1)。応答線については、トランスジーンの部位指向統合が、UAS調節遺伝子のゲノムコンテキストを固定するために、ΦC31触媒再結合カセット交換17,18によって行われるようになりました。このようにして、遺伝子挿入位置に関して遺伝子導入発現が正規化され、異なる候補遺伝子の表現型効果をより正確に比較することが可能になる。
これまでに作成された応答線は、高いレベルでトランスジーンを発現するか、RNA干渉(RNAi)を介して遺伝子発現を減少させるように設計されています。 通常、cDNAクローンはUAS配列に融合して適切な発現プラスミドを生成するが、完全なゲノム配列はクローニングには大きすぎるものではないと仮定しても可能である。サイレンシング構築物を生成するために、RNAiを刺激するヘアピンdsRNAを形成する適切なタンデム逆配列を得るために3つの異なる方法を用いた。これらには、融合PCR、非対称PCRおよびヘアピン構築物の商業的合成が含まれています。各方法に共通するのは、転写安定性を提供するために反転配列間にイントロン配列を含めることである。目的の遺伝子/RNAi構築物を挿入できる応答性プラスミドが開発された15。これらのプラスミドはまた、RMCE(RCME技術を詳細に記述するJoVE紙に付随するアドルフィに記載)に必要なΦC31 attB サイトを運びます。これらのプラスミドの1つに挿入するシーケンスを過剰発現にする際に必要な重要なステップをカバーするプロトコルが、この原稿に含まれている。また、RNAiヘアピン構築物作成用の2つのプロトコルを説明し、図示する。
新しいラインを作成する際には、まれなトランスジェニックな個体の同定は、トランスジェニックコロニーを確立し維持するために繁殖するために重要です。GAL4-UASシステムにとって最も重要なことは、応答者とドライバラインを区別して十字架を確立し、両方の遺伝子を運ぶ個々の子孫を特定する必要があります。これは、ドライバと応答側カセットにリンクされた異なる支配的な選択可能マーカー遺伝子を使用することによって達成される。最も一般的には、光学フィルター(例えば、eYFP、eCFP、dsRed)を使用して明確に識別可能な蛍光マーカー遺伝子です。マーカーは、異常や汚染の識別が容易になるため、既知の信頼性の高い時空間パターンで表現することが重要です。蛍光マーカー遺伝子発現は、合成 3xP3 プロモーターによって日常的に調節され、 An.gambiae 開発19のすべての段階で眼および腹側神経節特異的発現を引き起こす。 3xP3 で制御される蛍光マーカーは、本稿に記載されているすべての形質転換プラスミドに含まれています。蛍光 アンガンバエ PUPAe GAL4-UASラインをスクリーニングするために使用される一般的な方法を詳述するプロトコルがここに含まれています。
GAL4-UAS システムの重要な要素の 1 つは、差し切れにマークされたドライバとレスポンダラインを横断する必要があります。この男性と女性を各ラインから行うには、交配前に分離する必要があります。しかし、成人は視力によって容易に区別できるが、遺伝的十字架を確立するためには、交配が起こらないように成人の出現前に男女を分離することが賢明である。男性と女性の一般的なサイズの違い アンガンビアの 子犬は、性決定の効率的で信頼性の高い方法になるにはあまりにも可変です20。代わりに、外部生殖器の明確な形態学的違いは 、An.ガンビアでのセックスのための信頼できる基礎を提供します。この記事では、適切な十字架を設定するために An.ガンビア の子犬をセックスするための信頼できる方法について説明します。
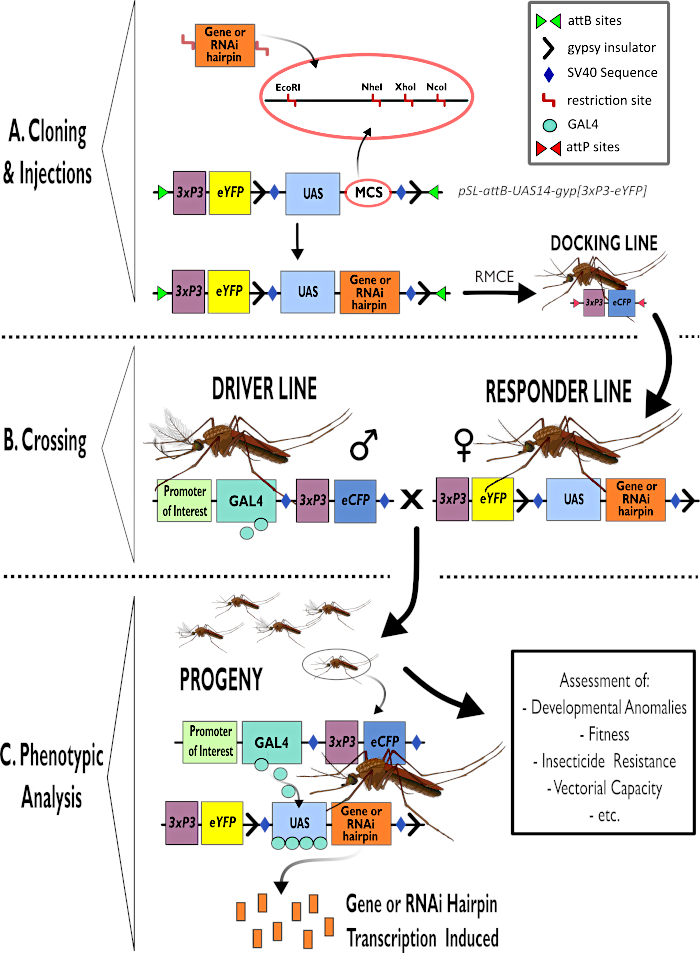
図1 -アノフェレスガンビアにおける二国間GAL4-UASシステムを使用するためのプロセスの図示的表現。 (A)例ベクトルの主要成分(pSL-attB-UAS14-gyp[3xp3-eYFP])が描かれ、対象遺伝子にヘアピン構造またはコード配列を挿入するのに適した複数のクローニング部位内の利用可能な制限部位(EcoRI、NheI、XhoIおよびNcoI)を詳述している。ドッキングラインの構造も描かれています。(B)交差工程は、ドライバーライン(3xP3プロモーターによって駆動される目的のプロモーターおよびeCFPによってGAL4ドライバーを運ぶ)および応答側ライン(UASプロモーターおよび3xP3プロモーターによって制御されるeYFPマーカーによって制御される関心のある遺伝子またはヘアピン構築物を運ぶ)からの雄の使用を示す図示される。(C)Bにおける十字架の子孫における目的の遺伝子のGAL4駆動発現の図示および評価される典型的な表現型の一部のリスト。略称:複数クローニング部位(MCS)、リコンビナーゼ媒介カセット交換(RMCE)、アップストリームアクチベーター配列(UAS)、強化された黄色蛍光タンパク質(eYFP)、強化シアン蛍光タンパク質(eCFP)。 この図の大きなバージョンを表示するには、ここをクリックしてください。
GAL4-UAS システムの二者間の性質を提供する十字の使用であり、より線形なアプローチよりも明確な利点があります。例えば、ドライバーとレスポンダーラインの組み合わせは、プロモーター/遺伝子の組み合わせごとに新しいトランスジェニックラインを生成して維持する必要がある場合よりも評価できます。さらに重要なことは、線形系で作成/維持することが困難な発現が摂動されたときに致死的または無菌表現型を産生する遺伝子の分析を可能にする。このような致死性表現型は、遺伝子機能および時空間的発現に応じて、すべての発達段階で現れるが、胚発生時に最も頻繁に観察される。蚊の胚の発達を視覚化するには、卵をコーティングする不透明な絨毛のクリアが必要です。Trpiš(1970)21 およびカイザーら(2014)22に記載されている方法に従って、我々は、構造的完全性を維持しながら、胚を固定するために使用するプロトコルを記述し、顕微鏡的可視化およびイメージングを可能にするエンドコリオンをクリアするために漂白する。
