1. QMC assembly
- Assemble QMCs from parts (Figure 1A) with a single egg laying plate (ELP) inserted as shown in Figure 1B. Do not add feeder tubes until after the workers have been added to the cage. Temporarily cover the 4 feeder holes with laboratory grade tape.
- Insert the queen excluder and the feeding chamber door over the feeding chamber to keep the queen from entering the feeding chamber and contacting the treated diet. See Fine et al.32 for further assembly details.
- Collect the wax comb frames containing the capped worker brood from honeybee colonies 24 h prior to adult eclosion and place them in an incubator (34.5 °C) inside a brood box. 24 h later, brush the eclosed bees off the frames and into an open container that has been lined with an insect barrier paint (e.g., Fluon) to prevent the bees from crawling out.
- Add at least 50 bees by weight (5 g ≈ 50 bees44,45) to the egg laying chamber of each QMC. To ensure that a diverse genetic pool of workers is represented in the experiment, obtain an approximately equal number of worker bees from at least three colonies and mix them prior to adding them to the QMCs.
NOTE: Newly eclosed worker bees less than 1 day old cannot fly or sting due to their underdeveloped flight muscles and unhardened cuticle. If they are added at this age, there is no need to anesthetize them prior to handling. They can be weighed by gently scooping bees from the container using a small ¼ cup volume measuring cup and placing them into a second container (lined with insect barrier paint e.g., Fluon) that has been tared on a scale. The area of the frames covered by capped brood should be roughly equal to ensure that source colonies are equally represented in the QMC worker populations. Homogenization of worker bees can be achieved by brushing newly eclosed bees from frames taken from all colonies into the same container and allowing them to mix for 5 min prior to adding them to QMCs. - Add the feeders containing sucrose solution, water, and pollen supplement (See section 2).
- Expose the individual mated queens to CO2 gas to stimulate egg laying46 and to ease transfer into QMC.
- Use queens purchased from a commercial breeder within 48 hours of receipt. While the queen is still inside the shipping cage, place it in a clear plastic bag. Place one end of a plastic tube connected to a CO2 gas cannister inside the bag and gently open the cannister valve to allow the CO2 gas to flow.
- When the bag has been inflated with gas, simultaneously close the cannister valve and hold the bag closed to trap the gas inside. Keep the bag closed for 30 s or until the queen has stopped moving. Remove the queen and open the shipment cage once she is observed to be unconscious.
- Partially open the door to the egg laying chamber, gently place the unconscious queen inside and close the lid, taking care not to crush the queen or workers inside. Add the second egg laying plate to each QMC as shown in Figure 1C. Place a piece of laboratory tape across the top of the two ELPs to keep them from separating from the QMC frame and prevent workers from exiting the cage.
- Place the cages in a dark incubator with stable environmental conditions of 34 ± 0.5 °C and 60% ± 10% relative humidity, like the conditions inside a normal colony.

Figure 1. A: Disassembled QMC. B: Partially assembled QMC with 1 ELP inserted. C: Fully assembled QMC with 2 ELPs. Please click here to view a larger version of this figure.
2. Preparing and administering diets laced with agrochemicals
- To prepare 1000 g of 50% (g/g) sucrose solution, place a stir bar in the bottom of a clean 1 L glass reagent bottle. Add 500 g sucrose and 500 mL of deionized water. Unscrew the lid of the bottle and use a heated stir plate set to low heat to mix the solution until all the sucrose has dissolved. Allow the solution to cool to room temperature before adding the agrochemical stock solutions.
- Prepare the stock solutions of agrochemicals in an appropriate solvent, such as acetone, at a concentration that can be added to diet to achieve the desired final concentration of the agrochemical of interest.
NOTE: When using acetone as a vehicle solvent, the Organization for Economic Cooperation and Development (OECD) guidelines stipulate that the final concentration of acetone in diet must be ≤ 5% for chronic oral toxicity tests on adult honey bees47. However, some solvents such as n-methyl-2-pyrrolidone5,31 and dimethyl sulfoxide25 can exert toxic effects below this concentration, so it is recommended to keep solvent concentrations as low as possible in treatment diet. Depending on the volume and type of solvent used, it may be necessary to include both a solvent control group and a negative control group to ensure that potential effects due to solvent toxicity are detected. When using formulated products, the amount of the product used must be adjusted based on the concentration present in the formulation. Depending on the stability of the agrochemical of interest in the solvent, stock solutions can be kept for up to 2 weeks at -20 °C. - Select sublethal doses based on the results of OECD Test No. 245: Honey Bee (Apis mellifera L.), Chronic Oral Toxicity Test (10-Day Feeding)47, and identify the relevant literature by querying the Ecotox knowledgebase48.
- Administer the agrochemical treatments in a sucrose solution, a commercial pollen supplement (if available as a powder), or both. Prepare the experimental diet for use the same day by adding an appropriate amount of stock solution to chilled/room temperature 50% sucrose solution (w/w). Mix thoroughly by vortexing or with a stir bar set to medium speed. For pollen supplements, add the agrochemical laced sucrose solution to the powdered supplement instead of the syrup according to manufacturer protocols, making sure to adjust the volume of stock solution used according to the final weight of the pollen diet. See Table 1 for example calculations.
- Prepare the feeder tubes from 2 mL microcentrifuge tubes.
- For liquid diet feeders, heat the tip of a 20-gauge needle on a hot plate/stovetop and puncture the bottom of the tube twice. Close the tube lid and pipette approximately 1.5 mL of sucrose solution or water through one of the puncture holes. Set the tube down with the punctured side up until it is added to the QMC.
- For pollen supplement feeders, use a razor blade to slice off the bottom of the tube. Close the lid and a push a 1-2 g ball of pollen supplement into the tube until it touches the lid.
- Record the feeder weights prior to placing them in the QMCs. Do not keep unused diet at 4 °C for over 48 hours.
| Desired Concentration | Desired Solvent Vehicle Concentration | Desired final volume/mass of sucrose solution | Voume of stock solution | Imidacloprid in stock solution | Suggested stock solution recipe | |
| Sucrose solution | 10 ppb (w/w) | 0.05% (v/v) | 81.45 mL/100 g* | 40.7 µL | 0.001 mg/40.7 µL | 0.02 mg/814 µL |
| Pollen supplement | 10 ppb (w/w) | 10 g** | 4.07 µL | 0.0001 mg/4.07 µL | 0.02 mg/814 µL | |
| Sucrose solution | 50 ppb (w/w) | 0.05% (v/v) | 78.5 mL/100 g* | 40.7 µL | 0.005 mg/40.7 µL | 0.1 mg/814 µL |
| Pollen supplement | 50 ppb (w/w) | 10 g** | 4.07 µL | 0.0005 mg/4.07 µL | 0.1 mg/814 µL |
Table 1: Example recipes for treated sucrose solution, pollen supplement, and stock solution. *Volume based on the density of 50% (w/w) sucrose solution (1.228 g/mL). **The density of the pollen supplement will vary depending on what product is used, but if this solvent volume is used, the final solvent concentration in pollen supplement will be within the desired range of ≤ 5% by volume.
3. Monitoring – Egg Production Rate
- Quantify the egg laying 1 to 2 times per day in the morning and/or evening. Begin by removing QMCs from the incubator to check for eggs.
NOTE: In a successful experiment, egg production will commence in most of the control QMCs within 3 days of initial cage assembly. Only take as many QMCs out of the incubator at one time that can be checked and fed within 10 min. Longer periods outside the incubator may disrupt egg production. - Examine the backs of the clear ELPs for eggs. If eggs are present, remove the door panel in front of the plate of interest. Remove the tape from across the ELPs and carefully slide the door panel between the ELP and the bees inside the QMC, taking care not to crush any bees that might be cleaning the cells in the ELPs.
- With the door panel in place, remove the ELP, and count and record the number of eggs inside the ELP cells. Remove the eggs by tapping the edge of the ELP, open cell-side down, on a hard surface (such as the lip of a waste receptacle). Once the eggs fall out, replace the empty ELP in the QMC. Gently remove and replace the door panel behind the ELP on the outside of the QMC. Repeat as necessary with the second ELP and replace the tape across the QMC when finished.
NOTE: Egg production generally declines and mortality increases in QMCs after 2 weeks32,33, therefore it is recommended to conclude experiments after 14 days.
4. Monitoring – Food Consumption
- Replace all the food remaining in QMC feeders with freshly prepared diet every two days. Prepare new feeder tubes (including water) and weigh them before removing QMCs from the incubator for monitoring. Swap all old tubes with new ones and weigh old tubes before disposing of unconsumed diet. Compare the final weight of the feeder tube and unconsumed diet to the weight of the same feeder tube prior to placing it in the QMC to estimate diet consumption.
- Between days when feeders are scheduled to be replaced, check diet consumption once per day (at the same time when QMCs are monitored for egg production) to ensure that feeders are never empty. If a feeder tube is empty or near empty, remove it, refill it, record the weight of the tube before and after and add the difference to the 2-day diet consumption total for the QMC.
5. Monitoring – Embryo Viability
- At a selected point during a QMC experiment, remove ELPs containing freshly laid eggs from the QMC according to step 3, but do not dislodge eggs from the ELP.
- Cover the ELP with a universal microplate lid and place it inside a desiccator with a saturated K2SO4 solution (150 g K2SO4 in 1 L of water, kept in a shallow dish).
NOTE: Some salt should be visible on the bottom of the dish after the mixture comes to temperature in the incubator. - Keep the desiccator in an incubator set to 34.5 °C, resulting in a relative humidity of 95% inside the desiccator, similar to the conditions used by Collins49.
NOTE: Almost all eggs will hatch within 72 ± 6 hours of when they were laid49, hence hatching rates can be assessed as early as 78 hours after the ELPs were removed from the QMC. A "C" shape larva in the bottom of the cell is indicative of a successful hatching event. Some variation in this timing is possible if, for example, the eggs are drones and not workers50.
6. Worker Sampling
- If the QMCs have been populated with excess workers, sample the worker bees at a selected time point during the experiment for assessment of treatment induced changes in their physiology. Perform the collections in conjunction with daily feeding and egg counting activities to minimize the time for which the QMCs are outside of the incubator.
- Before sampling, place a door panel between an ELP and the interior of the QMC, and remove the ELP. Carefully lift the door panel approximately 0.5 cm from the base of the cage and remove a worker bee from inside the QMC using featherweight tweezers. To prevent bees from escaping, cover portions of the 0.5 cm opening with a gloved finger or piece of cotton as necessary.
- Preserve the collected bee for follow-up analysis and repeat this process until the desired number of samples have been collected. For gene expression analysis, snap freezing bees in liquid nitrogen and immediate storage at -80 °C is strongly recommended51.
7. Worker Mortality
- Assess worker mortality during the experiment by counting the number of dead bees at the bottom of the feeding chamber and the egg laying chamber. Perform this assessment in conjunction with daily egg laying quantification.
- Using forceps, carefully remove the dead bees through the feeder holes, covering the hole with a gloved finger or piece of cotton while the forceps are not inserted.
- Remove the dead bees from the egg laying chamber by carefully lifting the door panel approximately 0.5 cm from the base of the cage and inserting forceps. To prevent bees from escaping, cover portions of the 0.5 cm opening with a gloved finger or piece of cotton as necessary.
- Assess the worker mortality at the conclusion of the experiment by removing and counting all the dead bees from the QMCs using the previously described methods prior to euthanizing the remaining bees.
NOTE: In the absence of worker bees, queens will not produce eggs and will starve within 24 hours. Therefore, if all the workers in a QMC are observed to be dead, the QMC should be removed from the experiment. Likewise, if a queen dies during the experiment, the QMC should be removed, and the data should be appropriately censored.
The production of eggs was monitored in QMCs assembled and maintained as described above with once daily observations of egg production and 15 cages per treatment group. Newly mated queens of primarily Carniolan stock were purchased and shipped overnight from a queen breeder, and honey bee workers were obtained from 3 colonies maintained according to standard commercial methods at The Bee Research Facility at the University of Illinois Urbana-Champaign. Here, 4 dietary treatment groups were used: 1) 50 ppb (g/g) imidacloprid in sucrose solution and pollen supplement (50 ppb – p+s), 2) 10 ppb imidacloprid in sucrose solution and pollen supplement (10 ppb – p+s), 3) 10 ppb imidacloprid in pollen supplement alone (10 ppb – p), and 4) a control group given diet containing an equivalent volume of acetone as the treatment groups (CTRL).
Treatment-related changes in daily egg counts were evaluated as described in Fine et al.32 with minor modifications. Briefly, a Poisson log-linear GEE with an auto-regressive (AR-1) correlation matrix structure was implemented to assess treatment related changes in egg production over time. Here, time (day) was treated as a continuous variable and treatment was categorical. Wald chi-square post hoc tests were used to determine significance. Because no egg laying was observed on day 1 of the experiment, this day was excluded from analysis to conform to the assumptions of the GEE. The results of this analysis are shown in Table S1. Daily egg production was significantly lower in QMCs in the 50 ppb p+s treatment group (χ2=43.99, p<0.001; Figure 2A).
Differences in the total number of eggs produced in QMCs by treatment were analyzed using a one-way ANOVA and Tukey HSD post hoc test (Figure 3). For this analysis, any QMC removed from the experiment before the end of the 14-day monitoring period due to queen or worker death was excluded, resulting in N=13 each for the CTRL and the 50 ppb – p+s groups, N=14 for 10 ppb – p, and N=15 for 10 ppb – p+s. A dose dependent effect was observed for treatments administered in both sucrose and pollen, with the largest reduction in egg production relative to control observed in 50 ppb – p+s followed by 10 ppb – p+s. No difference in total eggs produced was observed between CTRL and 10 ppb – p (F3, 52=17.95, p<0.001, Tukey HSD).
Consumption of pollen supplement and water was recorded every 48 hours for 10 days, and consumption of sucrose solution was recorded every 48 hours for 12 days. Changes in diet consumption rates were evaluated using Gaussian distributed GEEs with the same parameters as described above (Figure 2B-D). Results are summarized in Table S1. Briefly, daily rates of sucrose consumption significantly increased as the experiment progressed (χ2=6.03, p=0.014), but rates of pollen supplement consumption decreased (χ2=174.98, p<0.001). Significantly higher rates of pollen consumption were observed when imidacloprid was administered at 10 ppb in pollen supplement alone (χ2=21.44, p<0.001) and significantly decreased when it was administered at either 10 or 50 ppb in pollen supplement and sucrose solution together (10 ppb – p+s: χ2=6.59, p=0.010; 50 ppb – p+s: χ2=14.47, p=0.0001).
Eggs were collected from QMCs on day 7 of the experiment, and changes in the number of eggs hatching successfully following maternal exposure to agrochemical treatments was assessed using a generalized linear mixed model (GMLR) with a binomial distribution and QMC identity treated as a random effect. Maternal exposure to imidacloprid administered at 10 ppb in pollen alone or in pollen and sucrose solution did not affect egg hatching rates (10 ppb – p+s: Z=-0.139, p=0.290; 10 ppb – p: Z=0.182, p=0.856). Hatching rates could not be assessed for eggs laid by queens in QMCs provisioned with 50 ppb imidacloprid laced diet due to low rates of egg production in this treatment group.
For this work, all statistical analysis was performed in R Studio 1.2.5003 (Boston, MA, USA). Figures were prepared using JMP Pro 15 and Photoshop CC 2019 (Adobe Inc., San Jose, CA). Data are available in Supplementary file S1.
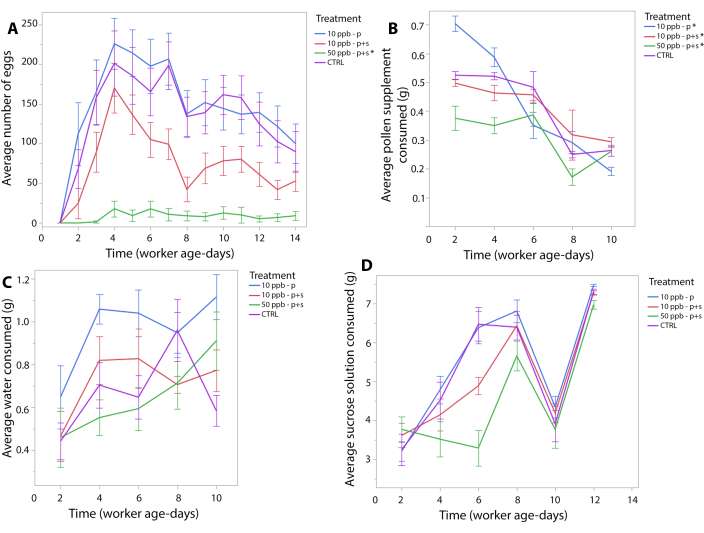
Figure 2. A: Average ± SE eggs per day in QMCs. B: Average ± SE pollen supplement, C: sucrose solution, D: and water (g) consumed during 48-hour periods in QMCs. Significance of treatments (indicated by "*") determined by GEE and Wald chi-square post hoc test. Please click here to view a larger version of this figure.
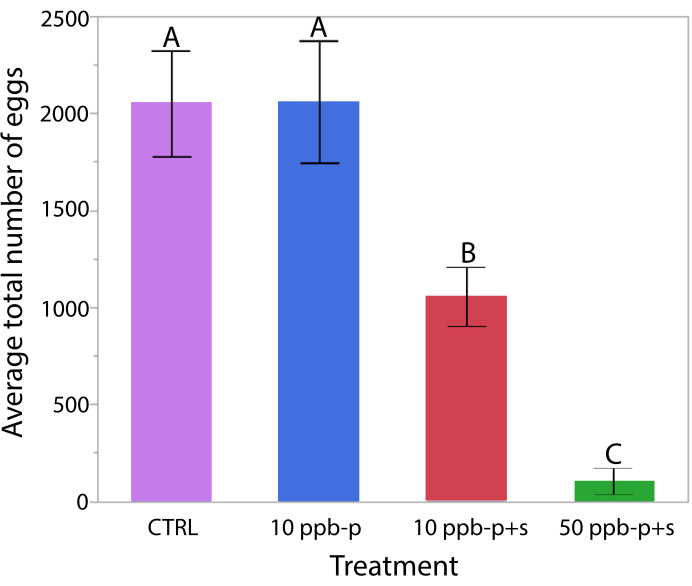
Figure 3: Average ± SE sum of eggs laid by treatment during experiment. Significance (indicated by letters) determined by ANOVA and Tukey HSD post hoc test. Please click here to view a larger version of this figure.
Table S1:Results of GEEs analyzing changes in egg laying rates and diet consumption in QMCs over time. Please click here to download this Table.
Supplementary file S1: Please click here to download this Supplemental File.
