Bacteriën gebruiken verschillende kleine signaalmoleculen om zich aan te passen aan constant veranderende omgevingen1,2. Bijvoorbeeld, de auto-inducers, N-acylhomoserine lactones en hun gemodificeerde oligopeptiden, bemiddelen de intercellulaire communicatie tussen bacteriën om populatiegedrag te coördineren, een fenomeen dat bekend staat als quorum sensing2. Een andere groep kleine signaalmoleculen zijn de NSM’s, waaronder het veel bestudeerde cyclische adenosinemonofosfaat (cAMP), cyclisch di-AMP, cyclisch di-guanosinemonofosfaat (cyclisch di-GMP) en guanosine penta- en tetrafosfaat (p)ppGpp1. Bacteriën produceren deze NSM’s als reactie op verschillende stressomstandigheden. Eenmaal geproduceerd, binden deze moleculen zich aan hun doeleiwitten en reguleren ze verschillende fysiologische en metabolische routes om de ondervonden stress aan te kunnen en de bacteriële overleving te verbeteren. Daarom is identificatie van de doeleiwitten een onvermijdelijke voorwaarde voor het ontcijferen van de moleculaire functies van deze kleine moleculen.
Het afgelopen decennium is getuige geweest van een hausse aan kennis van deze kleine signaalmoleculen, voornamelijk als gevolg van verschillende technische innovaties die de doeleiwitten van deze kleine moleculen onthulden. Deze omvatten de afvangverbindingstechniek3,4,5, en de differentiële radiale capillaire werking van ligandtest (DRaCALA)6 die in dit artikel moet worden besproken.
Uitgevonden door Vincent Lee en collega’s in 20116, DRaCALA zet het vermogen van een nitrocellulosemembraan in om differentieel vrije en eiwitgebonden liganden vast te zetten. Moleculen zoals eiwitten kunnen zich niet verspreiden op een nitrocellulosemembraan, terwijl kleine liganden, zoals de NSM’s, dat wel kunnen. Door het NSM (bijv.ppGpp ) te mengen met het te testen eiwit en deze op het membraan te spotten, kunnen twee scenario’s worden verwacht(figuur 1):Als (p)ppGpp zich aan het eiwit bindt, wordt het radiolabel (p)ppGpp in het midden van de vlek door het eiwit behouden en zal het niet naar buiten diffuus zijn, wat een intens klein puntje geeft (d.w.z. sterk radioactief signaal) onder een fosforimager. Als (p)ppGpp zich echter niet aan het eiwit bindt, zal het vrij naar buiten diffuus zijn om een grote vlek met een uniform radioactief achtergrondsignaal te produceren.
Bovendien kan DRaCALA de interactie tussen een klein molecuul en een ongezuiverd eiwit in een hele cellysaat detecteren als het eiwit in voldoende hoeveelheid aanwezig is. Deze eenvoud maakt het gebruik van DRaCALA mogelijk bij het snel identificeren van eiwitdoelen met behulp van een ORFeome-expressiebibliotheek. Doeleiwitten van cAMP7, cyclische di-AMP8, cyclische di-GMP9,10en (p)ppGpp 11,12,13 zijn systematisch geïdentificeerd met behulp van DRaCALA. Dit videoartikel gebruikt (p)ppGpp als voorbeeld om de kritische stappen en overwegingen bij het uitvoeren van een succesvolle DRaCALA-screening te demonstreren en te beschrijven. Een meer grondige beschrijving van DRaCALA14 wordt ten zeerste aanbevolen om te lezen in combinatie met dit artikel voordat U DRaCALA uitvoert.
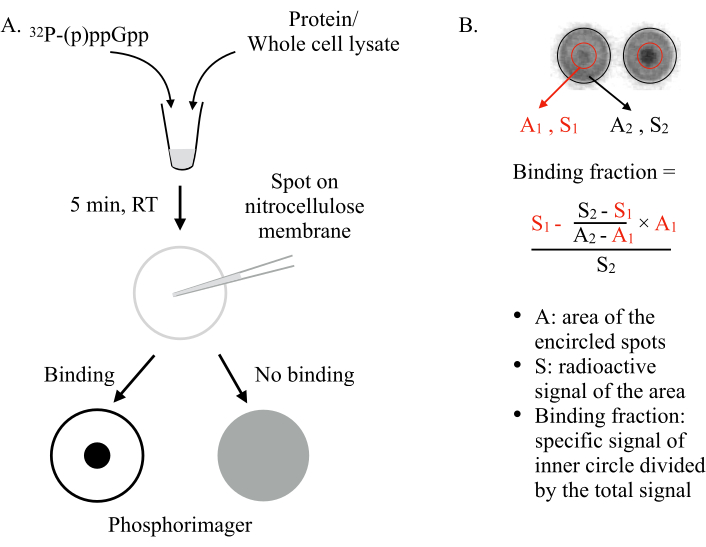
Figuur 1: Het principe van DRaCALA. (A) Schematisch van de DRaCALA-test. Zie de tekst voor meer informatie. B) Kwantificering en berekening van de bindende fractie. Zie de tekst voor meer informatie. Kortom, de DRaCALA-vlekken worden geanalyseerd door twee cirkels te tekenen die de hele vlek en de binnenste donkere stip omringen(d.w.z.de behouden (p)ppGpp vanwege de binding van het geteste eiwit). Het specifieke bindingssignaal is het radioactieve signaal van de binnenste cirkel (S1) na aftrek van het niet-specifieke achtergrondsignaal (berekend door A1 × ((S2-S1)/(A2-A1))). De bindingsfractie is het specifieke bindingssignaal gedeeld door het totale radioactieve signaal (S2). Afkortingen: DRaCALA = Differential Radial Capillary Action of Ligand Assay; (p)ppGpp = guanosine penta- en tetrafosfaten; RT = kamertemperatuur. Klik hier om een grotere versie van deze afbeelding te bekijken.
Het werk wordt ondersteund door een NNF-projectsubsidie (NNF19OC0058331) aan YEZ en het onderzoeks- en innovatieprogramma Horizon 2020 van de Europese Unie in het kader van de Marie Skłodowska-Curie-subsidieovereenkomst (Nº 801199) aan MLS.

