Bakterier bruker flere små signalmolekyler for å tilpasse seg stadig skiftende miljøer1,2. For eksempel, autoindusere, N-acylhomoserine laktoser og deres modifiserte oligopeptider, formidler intercellulær kommunikasjon blant bakterier for å koordinere befolkningsadferd, et fenomen kjent som quorum sensing2. En annen gruppe små signalmolekyler er NSM-ene, inkludert det mye studerte sykliske adenosinmonofosfatet (cAMP), syklisk di-AMP, syklisk di-guanosinmonofosfat (syklisk di-GMP) og guanosin penta- og tetrafosfater (p)ppGpp1. Bakterier produserer disse NSM-ene som et svar på en rekke ulike stressforhold. Når de er produsert, binder disse molekylene seg til sine målproteiner og regulerer flere forskjellige fysiologiske og metabolske veier for å takle de påløpkede påkjenningene og forbedre bakteriell overlevelse. Derfor er identifisering av målproteinene en uunngåelig forutsetning for å dechiffrere molekylære funksjoner til disse små molekylene.
Det siste tiåret har vært vitne til en boom av kunnskap om disse små signalmolekylene, hovedsakelig på grunn av flere tekniske innovasjoner som avduket målproteinene til disse små molekylene. Disse inkluderer fangstsammensetningsteknikken3,4,5og differensial radial kapillærvirkning av ligandanalyse (DRaCALA)6 som skal diskuteres i dette dokumentet.
Oppfunnet av Vincent Lee og medarbeidere i 20116, DRaCALA distribuerer evnen til en nitrocellulosemembran til differensialt sequester frie og proteinbundne ligander. Molekyler som proteiner kan ikke spre seg på en nitrocellulosemembran, mens små ligander, som NSM-ene, er i stand til det. Ved å blande NSM (f.eks.ppGpp) med proteinet som skal testes og oppdage dem på membranen, kan to scenarier forventes (Figur 1): Hvis (p)ppGpp binder seg til proteinet, vil den radiomerkede (p)ppGpp beholdes i midten av stedet av proteinet og vil ikke spre seg utover, noe som gir en intens liten prikk (dvs. radioaktivt signal) under en fosforimager. Men hvis (p)ppGpp ikke binder seg til proteinet, vil det spre seg fritt utover for å produsere et stort sted med ensartet bakgrunn radioaktivt signal.
Videre kan DRaCALA oppdage samspillet mellom et lite molekyl og et uoppgjort protein i en hel cellelyse hvis proteinet er tilstede i tilstrekkelig mengde. Denne enkelheten gjør det mulig å bruke DRaCALA i raskt å identifisere proteinmål ved hjelp av et ORFeome-uttrykksbibliotek. Faktisk er målproteiner av cAMP7, syklisk di-AMP8, syklisk di-GMP9,10og (p) ppGpp11,12,13 systematisk identifisert ved hjelp av DRaCALA. Denne videoartikkelen bruker (p)ppGpp som et eksempel for å demonstrere og beskrive de kritiske trinnene og vurderingene ved å utføre en vellykket DRaCALA-screening. Vær oppmerksom på at en grundigere beskrivelse av DRaCALA14 anbefales på det sterkeste å lese i kombinasjon med denne artikkelen før du utfører DRaCALA.
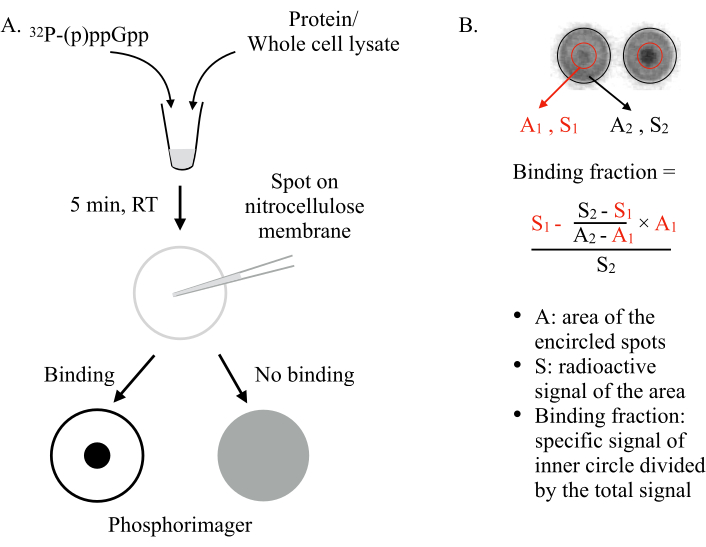
Figur 1: Prinsippet om DRaCALA. (A) Skjematisk for DRaCALA-analysen. Se teksten hvis du vil ha mer informasjon. (B) Kvantifisering og beregning av bindingsfraksjonen. Se teksten hvis du vil ha mer informasjon. Kort sagt vil DRaCALA-flekkene bli analysert ved å tegne to sirkler som omskriver hele stedet og den indre mørke prikken (dvs.beholdt (p) ppGpp på grunn av bindingen av det testede proteinet). Det spesifikke bindingssignalet er det radioaktive signalet til den indre sirkelen (S1) etter at det ikke-spesifikke bakgrunnssignalet er trukket fra (beregnet med A1 × ((S2-S1)/(A2-A1))). Bindingsfraksjonen er det spesifikke bindingssignalet delt på det totale radioaktive signalet (S2). Forkortelser: DRaCALA = Differensial radial kapillær virkning av Ligand Assay; (p)ppGpp = guanosin penta- og tetrafosfater; RT = romtemperatur. Klikk her for å se en større versjon av denne figuren.
Arbeidet støttes av et NNF Project Grant (NNF19OC0058331) til YEZ, og EUs forsknings- og innovasjonsprogram Horizon 2020 under Marie Skłodowska-Curie-tilskuddsavtalen (Nº 801199) til MLS.

