Pseudomonas aeruginosa (Pa) es un patógeno oportunista que establece infecciones crónicas en individuos inmunocomprometidos. Para aquellos con la enfermedad genética fibrosis quística (FQ), estas infecciones pueden abarcar el curso de toda la vida. La FQ causa la acumulación de un esputo viscoso y rico en nutrientes en las vías respiratorias, que se coloniza por una variedad de patógenos microbianos con el tiempo. Pa es uno de los patógenos de la FQ más prevalentes, colonizando las vías respiratorias en la primera infancia y estableciendo infecciones difíciles de tratar1. La PA sigue siendo un problema clínico significativo y se considera una de las principales causas de mortalidad en las personas con FQ, a pesar de la mejora de los regímenes terapéuticos en los últimos años2,3. Este fenotipo de persistencia y el aumento de la tolerancia a los antibióticos le han valido a Pa un lugar en un grupo de patógenos identificados tanto por los Centros para el Control de Enfermedades (CDC) como por la Organización Mundial de la Salud (OMS) como prioridades de investigación para el desarrollo de nuevas estrategias terapéuticas: los patógenos ESKAPE4.
Al igual que otros patógenos ESKAPE, la resistencia a los antibióticos adquirida es común en Pa,pero también hay muchas propiedades intrínsecas que contribuyen a la tolerancia antimicrobiana de Pa. Entre estos se encuentra la capacidad de Pa para formar agregados, grupos altamente densos de ~ 10-1,000 células, que se pueden observar en múltiples infecciones, incluido el esputo del paciente con FQ5,6. Al igual que el Pa estudiado en otros sistemas de biopelículas, los agregados de Pa muestran fenotipos clínicamente relevantes, como el aumento de la resistencia a los antibióticos y la activación de la comunicación célula-célula (detección de quórum (QS)). Por ejemplo, se ha demostrado que los agregados de Pa utilizan comportamientos regulados por QS para combatir otros microbios, así como tolerar tratamientos antimicrobianos como la producción de piocianina7. La capacidad de estudiar tales comportamientos ofrece una visión emocionante de los ecosistemas bacterianos en un entorno similar a aquel en el que existen en el cuerpo humano.
Uno de los mayores desafíos para estudiar cómo los agregados de Pa responden al entorno cambiante del esputo es la falta de sistemas nutricionalmente relevantes y robustos que promuevan la formación de agregados. Gran parte de lo que se sabe sobre Pa se ha descubierto utilizando sistemas in vitro en los que las células crecen planctonicamente o en una arquitectura característica de “hongo” unida a la superficie que no se ha observado in vivo8. Si bien los modelos clásicos de crecimiento de biopelículas, como las células de flujo o el agar sólido, han producido un conocimiento extenso y valioso sobre los comportamientos bacterianos y los mecanismos de tolerancia a los antibióticos, estos hallazgos no siempre se traducen in vivo. Muchos modelos in vitro tienen una capacidad limitada para imitar el entorno de crecimiento del sitio de infección humana, lo que requiere costosos estudios in vivo. A su vez, muchos modelos in vivo carecen de la flexibilidad y resolución que ofrecen las técnicas in vitro.
El esputo sintético de fibrosis quística (SCFM2) está diseñado para proporcionar un entorno para el crecimiento de Pa similar al experimentado durante la infección crónica en el pulmón de la FQ. SCFM2 incluye fuentes nutricionales identificadas en la sputa de FQ expectorada, además de mucina, lípidos y ADN. El crecimiento de Pa en SCFM2 requiere un conjunto de genes casi idéntico al requerido para el crecimiento en esputo real y apoya la formación natural de agregados de Pa 9,10. Después de la inoculación, las células planctónicas forman agregados que aumentan de tamaño a través de la expansión. Las células individuales (denominadas migrantes) se liberan de los agregados, migran a áreas no coloreadas y forman nuevos agregados10. Esta historia de vida se puede observar utilizando CLSM y análisis de imágenes a la resolución de una sola célula. Los agregados de Pa formados en SCFM2 son de tamaños similares a los observados en la FQ pulmonar10. Este modelo permite la observación de múltiples agregados de tamaño variable en tiempo real y en tres dimensiones a escala de micras. La microscopía de lapso de tiempo permite el seguimiento de miles (~ 50,000) de agregados en un experimento. El uso de software de análisis de imágenes permite la cuantificación de fenotipos agregados a partir de micrografías, incluido el volumen agregado, el área de superficie y la posición en tres dimensiones hasta los 0,1 μm más cercanos, tanto a nivel de agregado individual como de población. Tener la capacidad de agrupar agregados por fenotipo y posición permite la diferenciación de agregados en diferentes etapas de desarrollo con precisión, así como su respuesta a un microambiente cambiante6,11.
La aplicación de SCFM2 para estudiar agregados de Pa en ensayos de bajo volumen y alto rendimiento lo convierten en un modelo flexible y rentable. Como medio definido, SCFM2 ofrece uniformidad y reproducibilidad a través de múltiples plataformas, proporcionando un método nutricional y físicamente relevante para estudiar agregados de Pa in vitro9. Las aplicaciones incluyen su uso en combinación con CLSM para observar la organización espacial y la tolerancia a los antibióticos a alta resolución (como se describe en este documento de métodos). La capacidad de realizar experimentos que proporcionan datos en tiempo real a escala de micras permite el estudio de las interacciones intra-especie e inter-especie a medida que pueden ocurrir in vivo. Por ejemplo, SCFM2 se ha utilizado previamente para estudiar la dinámica espacial de la comunicación célula-célula en poblaciones agregadas a través de una red de sistemas utilizados por Pa para regular múltiples genes que contribuyen a la virulencia y patogénesis6.
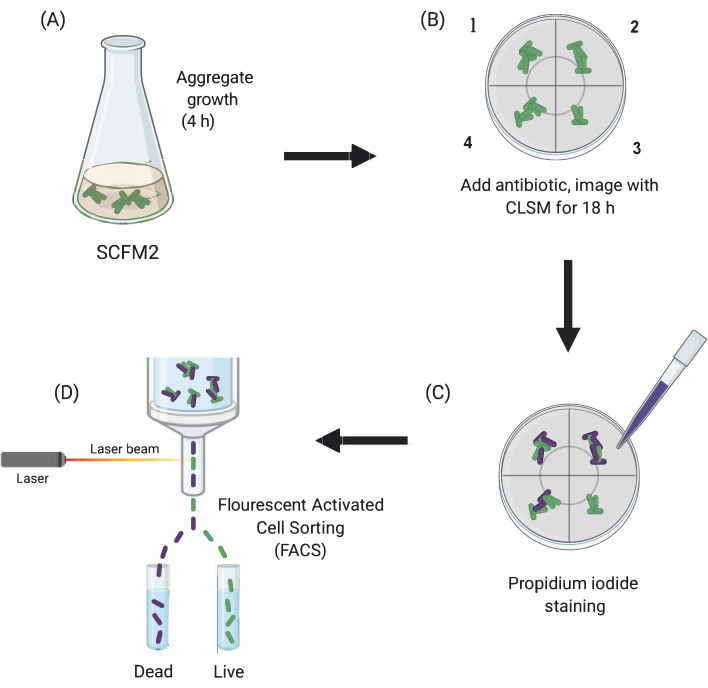
Figura 1: Representación gráfica de los principales pasos experimentales. (A) SCFM2 se inocula con células Pa y se le permite formar agregados en un plato de cultivo con fondo de vidrio. (B) Los agregados se transfieren al microscopio confocal y se agrega antibiótico. Se representan tres réplicas técnicas (cámaras 1-3) y un pozo de control (4) de SCFM2 inoculado sin tratamiento antibiótico. Los agregados se visualizan utilizando CLSM en el transcurso de 18 h. (C) Después de la imagen inicial de 18 h, los agregados se tratan con yoduro de propidio para visualizar las células muertas y se visualizan utilizando CLSM (D) Los agregados con el fenotipo deseado se separan de SCFM2 utilizando FACS. Abreviaturas: SCFM2 = medio de esputo sintético de fibrosis quística; Pa = Pseudomonas aeruginosa; CLSM = microscopía de barrido láser confocal; FACS = clasificación celular activada por fluorescencia. Haga clic aquí para ver una versión más grande de esta figura.
Aquí, se demuestra la utilidad de SCFM2 para estudiar el impacto del tratamiento antibiótico en los agregados de Pa en tiempo real, seguido por el uso de un enfoque de clasificación celular para aislar poblaciones de agregados con fenotipos distintos para el análisis posterior(Figura 1).
| Amino acids | |||
| Alanine | Acr s Organics s Organics |
56-41-7 | |
| Arginine HCl | MP | 1119-34-2 | |
| Asparagine | Acrs Organics |
56-84-8 | Prepared in 0.5 M NaOH |
| Cystine HCl | Alfa Aesar | L06328 | |
| Glutamic acid HCl | Acrs Organics |
138-15-8 | |
| Glycine | Acrs Organics |
56-40-6 | |
| Histidine HCl H2O | Alfa Aesar | A17627 | |
| Isoleucine | Acrs Organics |
73-32-5 | |
| Leucine | Alfa Aesar | A12311 | |
| Lysine HCl | Alfa Aesar | J62099 | |
| Methionine | Acrs Organics |
63-68-3 | |
| Ornithine HCl | Alfa Aesar | A12111 | |
| Phenylalanine | Acrs Organics |
63-91-2 | |
| Proline | Alfa Aesar | A10199 | |
| Serine | Alfa Aesar | A11179 | |
| Threonine | Acrs Organics |
72-19-5 | |
| Tryptophan | Acrs Organics |
73-22-3 | Prepared in 0.2 M NaOH |
| Tyrosine | Alfa Aesar | A11141 | Prepared in 1.0 M NaOH |
| Valine | Acrs Organics |
72-18-4 | |
| Antibiotic | |||
| Carbenicillin | Alfa Aesar | J6194903 | |
| Day-of Stocks | |||
| CaCl2 * 2H2O | Fisher Chemical | C79-500 | |
| Dextrose (D-glucose) | Fisher Chemical | 50-99-7 | |
| 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC) | Fisher (Avanti Polar Lipids) | 4235-95-4 | shake 15-20 min at 37 °C to evaporate chloroform |
| FeSO4 * 7H2O | Acrs Organics |
7782-63-0 | this stock equals 1 mg/mL, MUST make fresh |
| L-lactic acid | Alfa Aesar | L13242 | pH stock to 7 with NaOH |
| MgCl2 * 6H2O | Acrs Organics |
7791-18-6 | |
| N-acetylglucosamine | TCI | A0092 | |
| Prepared solids | |||
| Porcine mucin | Sigma | M1778-100G | UV-sterilize |
| Salmon sperm DNA | Invitrogen | 15632-011 | |
| Stain | |||
| Propidium iodide | Alfa Aesar | J66764MC | |
| Salts | |||
| K2SO4 | Alfa Aesar | A13975 | |
| KCl | Alfa Aesar | J64189 | add solid directly to buffered base |
| KNO3 | Acrs Organics |
7757-79-1 | |
| MOPS | Alfa Aesar | A12914 | add solid directly to buffered base |
| NaCl | Fisher Chemical | S271-500 | add solid directly to buffered base |
| Na2HPO4 | RPI | S23100-500.0 | |
| NaH2PO4 | RPI | S23120-500.0 | |
| NH4Cl | Acrs Organics |
12125-02-9 | add solid directly to buffered base |
| Consumables | |||
| Conical tubes (15 mL) | Olympus plastics | 28-101 | |
| Conical tubes (50 mL) | Olympus plastics | 28-106 | |
| Culture tubes w/air flow cap | Olympus plastics | 21-129 | |
| 35 mm four chamber glass-bottom dish | CellVis | NC0600518 | |
| Luria Bertani (LB) broth | Genessee Scientific | 11-118 | |
| Phosphate-buffered saline (PBS) | Fisher Bioreagents | BP2944100 | |
| Pipet tips (p200) | Olympus plastics | 23-150RL | |
| Pipet tips (p1000) | Olympus plastics | 23-165RL | |
| Serological pipets (5 mL) | Olympus plastics | 12-102 | |
| Serological pipets (25 mL) | Olympus plastics | 12-106 | |
| Serological pipets (50 mL) | Olympus plastics | 12-107 | |
| Ultrapure water (RNAse/DNAse free); nanopure water | Genessee Scientific | 18-194 | Nanopure water used for preparation of solutions in Table 1 |
| Syringes (10 mL) | BD | 794412 | |
| Syringes (50 mL) | BD | 309653 | |
| 0.22 mm PES syringe filter | Olympus plastics | 25-244 | |
| PS cuvette semi-mico | Olympus plastics | 91-408 | |
| Software | |||
| Biorender | To prepare the figures | ||
| FacsDiva6.1.3 | Becton Dickinson, San Jose, CA | ||
| Imaris | Bitplane | version 9.6 | |
| Zen Black | |||
| Equipment | |||
| FacsAriallu | Becton Dickinson, San Jose, CA | ||
| LSM 880 confocal laser scanning microscope | Zeiss |
