几年来,嗜血性卵母细胞已成为研究核迁移的示范系统。嗜血性卵母细胞在称为卵室的多细胞结构中发育。卵室包括16个生殖细胞(15个护士细胞和卵母细胞),周围环绕着上皮层的卵泡体细胞。卵室发育被细分为14个阶段(图1A),在此期间卵母细胞将生长并积累胚胎早期发育所需的储备。在开发过程中,在微管重组和母体决定因素的不对称传输时,卵母细胞沿着前体和多鼻体轴两极分化。这些轴确定胚胎和成人的后续极性轴产生于这个卵母细胞1的受精。在形成过程中,细胞核在卵母细胞中采用不对称位置。在第6阶段,细胞核以细胞为中心。当卵母细胞接收到的后卵泡细胞发出的尚未识别的信号时,细胞核会在第7阶段(图1B)2、3阶段向前等离子体膜和横向等离子体膜之间的交叉处迁移。此不对称位置需要诱导多索-文特轴的确定。
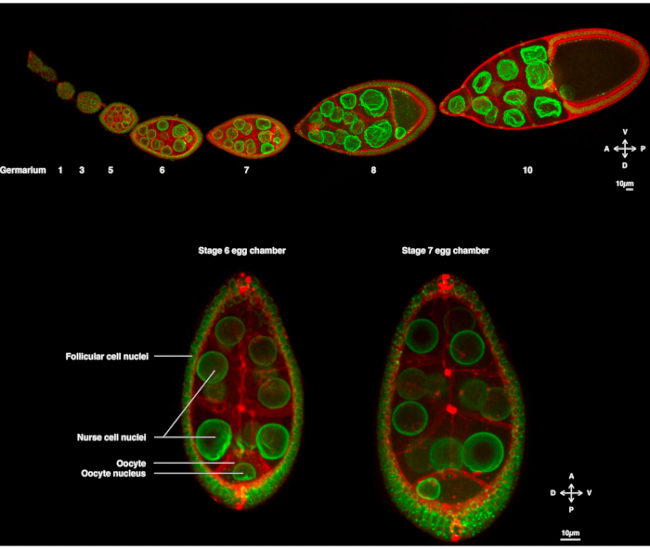
图1:果蝇黑色素加斯特卵室。(A) 转基因苍蝇的固定卵磷表示Fs(2)Ket-GFP,标记核信封和泛素-PH-RFP,标记等离子体膜。卵变由不同阶段的卵子室组成。成熟度沿着前后轴增加,前尖端(左)的天体和后尖的较旧阶段(右)。(B) 活蛋室的Z-投影,通过旋转盘共聚焦显微镜在卵母细胞的6级(左),其中细胞核以卵母细胞为中心。核将迁移到第7阶段(右)与前血浆膜(卵母细胞和护士细胞之间)和侧向血浆膜(卵母细胞和卵泡细胞之间)接触时采用不对称位置。此位置将诱导经侧的确定,从而诱导蛋室的圆周- 静脉轴。请单击此处查看此图的较大版本。
几十年来,这种核迁移一直通过免疫染色在固定组织上进行研究。这种方法显然证明这个过程依赖于一个密集的微管网络4,5。最近,我们开发了一个协议,提供条件兼容的活成像的卵母细胞在几个小时内,使有可能研究这个过程动态6。
因此,我们首次能够描述,细胞核在迁移过程中具有优先和特征轨迹,一个沿着前等离子膜(APM),另一个沿着卵母细胞的横向等离子膜(LPM)(图2)。这些最新成果强调了实时成像协议在研究核迁移等动态过程时的重要性。
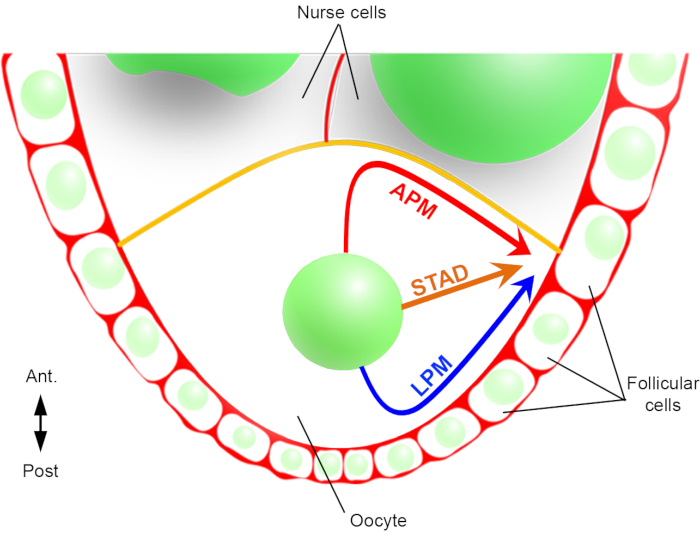
图2:细胞核不同迁移路径的示意图表示。在卵母细胞形成的第6阶段,卵母细胞是一个具有中央核的大细胞。在此阶段,前后极性轴设置与卵母细胞的后/横等离子膜接触卵泡细胞和前血浆膜(黄色)与护士细胞2接触。我们以前曾报告说,细胞核可以沿着前等离子膜(APM),沿着侧质等离子膜(LPM),或通过细胞质(STAD,直接迁移到前额皮层)6。请单击此处查看此图的较大版本。
卵母细胞核迁移是一种大约3h6的现象,到目前为止,触发实际迁移开始的事件还不得而知。用于研究这种机制的蛋白质突变体也会推迟迁移的开始。这些未知变量促使我们获得图像在很长一段时间内(10-12小时)。因此,确保卵母细胞存活是很重要的。随着蛋室的发展,它沿着前后轴从球形拉长到椭圆形。这种拉长是由卵泡细胞的旋转驱动的,卵泡细胞从第1阶段到第8阶段,垂直于前后轴7。此外,卵室周围还环绕着具有脉动特性肌肉的管状护套。其生理功能是将发育中的卵泡连续推向卵管8。为了限制解剖后诱导卵室振荡的运动,我们设计了一个高度为150微米的观测微室(图3A)。这个高度略高于10级和11级的卵泡大小。它大大限制了样品的垂直运动,同时保留了卵室的旋转,从而导致卵泡发育的有限缺陷。然后,我们使用旋转盘共聚焦显微镜对解剖的卵室进行 12 小时的实时成像。在这里,我们描述了我们研究第6阶段和第7阶段之间的卵母细胞核迁移的议定书。
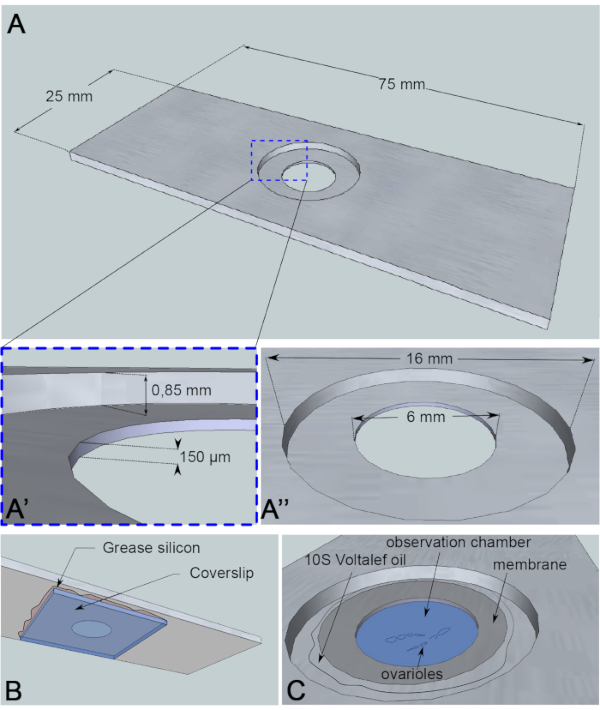
图3:观察室的示意图表示。(A) (上视图) 铝滑梯的精确尺寸与在滑梯中间钻的井的高度 (A’) 和周长 (A’) 。(B) (下视图) 封堵油井的盖片用硅润滑脂密封在滑梯上。(C) (上图) 解剖的卵巢在气体渗透膜覆盖的成像介质中发展。光环碳油用于稳定膜。请单击此处查看此图的较大版本。
为了跟踪核迁移并精确评估卵母细胞的轨迹,需要核包络和等离子体膜的标记。为此,选择了两个信号/噪声比高且不会在实时成像过程中褪色的转基因基因。为了标记等离子膜,建议使用 P[ubi-PH-RFP] 对人类磷脂酶C ∂(PLC∂1)的普莱克斯特林同源(PH)域进行编码。此 PH 域与沿卵母细胞9的等离子膜分布的磷酸二肽 PI (4,5)P2 结合。对于核包络 , P [ PPT – un1 ] Fs ( 2 ) Ket – GFP 蛋白陷阱菌株,其中 GFP 入编码的基因中 , Drosophila+ – 进口显示一个同质和强烈的信号10 。幼蝇(1-2天大)在卵巢解剖前被放置在含有干酵母24-48小时的新鲜小瓶中。
对于这种实时成像检测,一块1毫米厚的铝片,对样品没有反应,已被切成显微镜滑梯的尺寸。滑梯中央有一个直径为16毫米的孔,该孔已被反向到0.85毫米。这个计数器有一个直径为6毫米的孔,深度为150μm(图3A)。盖片粘在铝室底部的硅胶油脂(样品惰性)(图3B)。将样品放入中等填充井中后,将渗透到O2/CO2交换的膜放在中型油上,周围是卤化碳油(图3C)。
对于解剖,建议使用尖端尺寸为 0.05 x 0.02 mm 的不锈钢钳子和直径为 0.20 mm 的针头进行卵巢分离(图 4B,C)。迁移核在装有摄像头的旋转盘反向显微镜 CSU-X1 上成像。在 24 °C 下每 15 分钟通过延时获取多位置图像。 15 分钟的间隔允许进行多位置采集,对荧光蛋白的光照漂烁有限,并且样品的光毒性有限。此外,较短的间隔不会提供更多的信息数据来跟踪核轨迹。电影是通过斐济软件11处理和分析。
