Depuis plusieurs années, l’ovocyte de la drosophile est devenu un système modèle pour étudier la migration nucléaire. L’ovocyte de la drosophile se développe dans une structure multicellulaire appelée chambre à œufs. Les chambres à ovules englobent 16 cellules germinales (15 cellules nourricières et l’ovocyte) entourées d’une couche épithéliale de cellules somatiques folliculaires. Le développement de la chambre à ovules a été subdivisé en 14 étapes(Figure 1A),au cours desquelles l’ovocyte se développera et accumulera les réserves nécessaires au développement précoce de l’embryon. Au cours du développement, lors de la réorganisation des microtubules et du transport asymétrique des déterminants maternels, l’ovocyte se polarise le long des axes antéro-dorsal et dorso-ventral. Ces axes déterminent les axes de polarité ultérieurs de l’embryon et de l’adulte résultant de la fécondation de cet ovocyte1. Au cours de l’oogenèse, le noyau adopte une position asymétrique dans l’ovocyte. Au stade 6, le noyau est centré dans la cellule. Sur un signal encore à identifier émis par les cellules folliculaires postérieures qui est reçu par l’ovocyte, le noyau migre vers l’intersection entre les membranes plasmiques antérieure et latérale au stade 7 (Figure 1B)2,3. Cette position asymétrique est nécessaire pour induire la détermination de l’axe dorso-ventral.
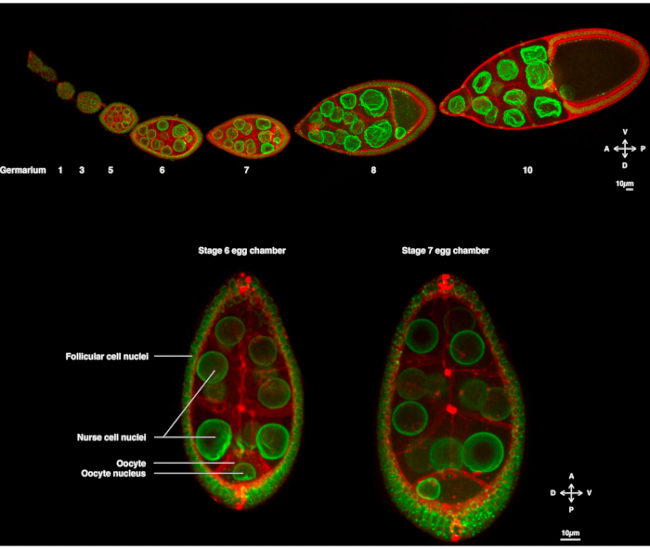
Figure 1: Chambres à œufs de Drosophila melanogaster. (A) Ovariole fixe de mouches transgéniques exprimant Fs(2)Ket-GFP qui marque les enveloppes nucléaires et ubi-PH-RFP qui marque les membranes plasmiques. L’ovariole est composée de chambres à œufs en développement à différents stades. La maturation augmente le long de l’axe antéro-postérieur avec le germarium à l’extrémité antérieure (à gauche) où réside la cellule souche germinale et le stade plus ancien à l’extrémité postérieure (à droite). (B) Projection Z de la chambre à œufs vivante par microscopie confocale à disque rotatif au stade 6 de l’oogenèse (à gauche), dans laquelle le noyau est centré dans l’ovocyte. Le noyau migrera pour adopter une position asymétrique au stade 7 (à droite) au contact de la membrane plasmique antérieure (entre l’ovocyte et la cellule nourricière) et de la membrane plasmique latérale (entre l’ovocyte et les cellules folliculaires). Cette position induira la détermination de la face dorsale et, par conséquent, de l’axe dorso-ventral de la chambre à œufs. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Pendant de nombreuses décennies, cette migration nucléaire a été étudiée sur des tissus fixes par immunocoloration. Cette approche a notamment permis de démontrer que ce procédé dépend d’un réseau dense de microtubules4,5. Plus récemment, nous avons développé un protocole proposant des conditions compatibles avec l’imagerie en direct de l’ovocyte pendant plusieurs heures permettant d’étudier ce processus dynamiquement6.
Ainsi, pour la première fois, nous avons pu décrire que le noyau a des trajectoires préférentielles et caractéristiques lors de sa migration, l’une le long de la membrane plasmique antérieure (APM) et l’autre le long de la membrane plasmique latérale (LPM) de l’ovocyte(Figure 2). Ces derniers résultats soulignent l’importance des protocoles d’imagerie en direct lors de l’étude des processus dynamiques tels que la migration nucléaire.
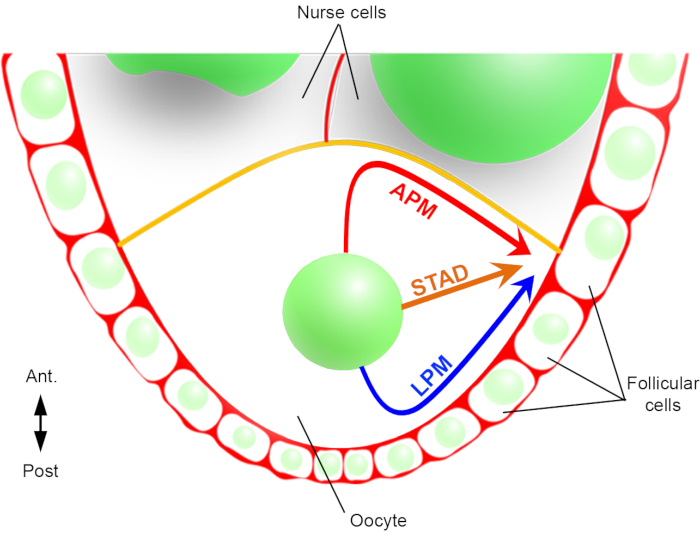
Figure 2: Représentation schématique des différentes voies de migration du noyau. Au stade 6 de l’oogenèse, l’ovocyte est une grande cellule avec un noyau central. A ce stade, l’axe de polarité antéro-postérieur est fixé avec une membrane plasmique postérieure/latérale de l’ovocyte en contact avec les cellules folliculaires et la membrane plasmique antérieure (en jaune) est en contact avec les cellules nourricières2. Nous avons précédemment rapporté que le noyau pouvait migrer soit le long de la membrane plasmique antérieure (APM), le long de la membrane plasmique latérale (LPM), soit à travers le cytoplasme (STAD, directement vers le cortex antéro-dorsal)6. Veuillez cliquer ici pour voir une version agrandie de cette figure.
La migration du noyau ovocytaire est un phénomène d’environ 3 h6, et jusqu’à présent, l’événement déclenchant le début de la migration réelle est inconnu. Le début de la migration peut également être retardé par les mutants protéiques utilisés pour étudier ce mécanisme. Ces variables inconnues nous ont motivés à acquérir des images sur de longues périodes (10-12 h). Il est donc important de s’assurer que les ovocytes restent vivants. Au fur et à mesure que la chambre à œufs se développe, elle s’allonge le long de l’axe antéro-postérieur d’une forme sphérique à une forme elliptique. Cet allongement est entraîné par la rotation des cellules folliculaires, qui se produit du stade 1 au stade 8, perpendiculairement à l’axe antéro-postérieur7. De plus, une gaine tubulaire de muscle à propriété pulsatile entoure les chambres à œufs. Sa fonction physiologique est de pousser les follicules en développement vers l’oviducte en continu8. Afin de limiter les mouvements qui induisent des oscillations des chambres à œufs après leur dissection, nous avons conçu une micro-chambre d’observation mesurant 150 μm de hauteur(Figure 3A). Cette hauteur est légèrement supérieure à la taille d’un follicule aux stades 10 et 11. Il limite considérablement les mouvements verticaux de l’échantillon tout en préservant la rotation de la chambre à œufs, ce qui entraîne des défauts limités dans le développement des follicules. Nous effectuons ensuite une imagerie en direct pendant 12 h sur des chambres à œufs disséquées par des acquisitions time-lapse multi-positions à l’aide d’un microscope confocal à disque rotatif. Nous décrivons ici notre protocole d’étude de la migration nucléaire des ovocytes entre les stades 6 et 7.
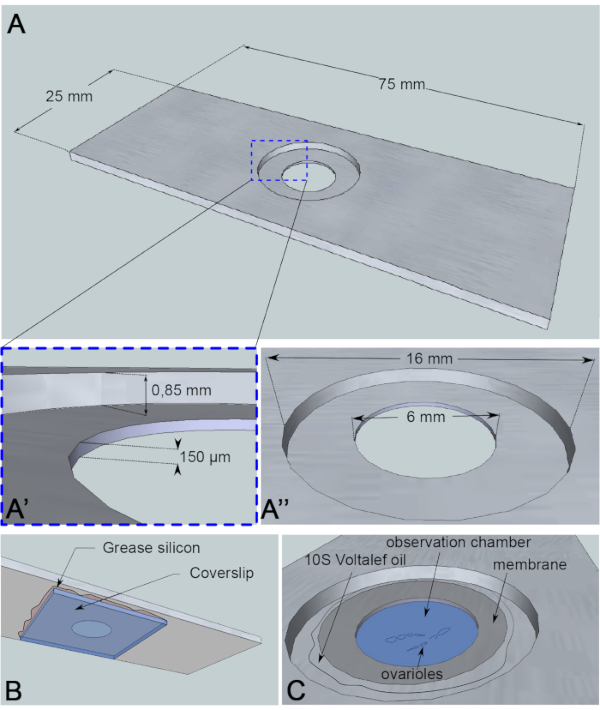
Figure 3: Représentation schématique de la chambre d’observation. (A) (Vue de dessus) Dimensions précises de la glissière en aluminium avec les hauteurs (A’) et les circonférences (A”) du puits foré au milieu de la glissière. (B) (Vue du bas) Un couvercle bloquant le puits est scellé à la glissière avec de la graisse de silicium. (C) (Vue de dessus) Les ovarioles disséqués se développent dans un milieu d’imagerie recouvert d’une membrane perméable aux gaz. L’huile d’halocarbure est utilisée pour stabiliser la membrane. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Afin de suivre la migration nucléaire et d’évaluer avec précision les trajectoires dans l’ovocyte, des marqueurs pour l’enveloppe nucléaire et la membrane plasmique sont nécessaires. Dans ce but, deux transgènes ayant un rapport signal/bruit élevé et ne s’estompant pas au cours de l’imagerie en direct ont été sélectionnés. Pour étiqueter la membrane plasmique, l’utilisation d’un P[ubi-PH-RFP] qui code le domaine d’homologie de Pleckstrin (PH) de la phospholipase humaine C ∂1 (PLC∂1) fusionnée à la RFP est recommandée. Ce domaine PH se lie au phosphoinositide PI(4,5)P2 distribué le long de la membrane plasmique de l’ovocyte9. Pour l’enveloppe nucléaire, la souche piège protéique P[PPT-un1]Fs(2)Ket-GFP où la GFP est insérée dans le gène codant pour la drosophile ß-importine présente un signal homogène et intense10. Les jeunes mouches (âgées de 1 à 2 jours) sont placées dans des flacons frais contenant de la levure sèche 24 à 48 heures avant la dissection des ovaires.
Pour ce test d’imagerie en direct, un morceau d’aluminium de 1 mm d’épaisseur, qui n’est pas réactif pour l’échantillon, a été découpé dans les dimensions d’une lame de microscopie. Il a un trou de 16 mm de diamètre au centre de la glissière qui a été contre-alésé à 0,85 mm. Ce contre-alésage a un trou supplémentaire de 6 mm de diamètre avec une profondeur de 150 μm(Figure 3A). Un couvercle est collé avec de la graisse de silicone (inerte pour l’échantillon) au fond de la chambre en aluminium(Figure 3B). Après avoir placé les échantillons dans le puits rempli de milieu, une membrane perméable à l’échange O2/CO2 est placée sur le milieu et entourée d’huile halocarbonée(Figure 3C).
Pour la dissection, il est recommandé d’utiliser des pinces en acier inoxydable d’une dimension de pointe de 0,05 x 0,02 mm et des aiguilles de 0,20 mm de diamètre pour la séparation des ovarioles (Figure 4B, C). Les noyaux migrateurs sont imagés sur un microscope confocal inversé à disque rotatif CSU-X1 équipé d’une caméra. Les images multi-positions ont été acquises par time-lapse toutes les 15 minutes à 24 °C. Un intervalle de 15 minutes permet d’effectuer des acquisitions multi-positions avec un photobleachage limité des protéines fluorescentes et une phototoxicité pour les échantillons. En outre, un intervalle plus court ne fournirait pas beaucoup plus de données informatives pour suivre les trajectoires nucléaires. Les films sont traités et analysés via le logiciel Fidji11.
