Per diversi anni, l’ovocita Drosophila è emerso come un sistema modello per studiare la migrazione nucleare. L’ovocita Drosophila si sviluppa in una struttura multicellulare chiamata camera dell’uovo. Le camere degli ovuli comprendono 16 cellule germinali (15 cellule nutrici e l’ovocita) circondate da uno strato epiteliale di cellule somatiche follicolari. Lo sviluppo della camera d’uovo è stato suddiviso in 14 fasi (Figura 1A), durante le quali l’ovocita crescerà e accumulerà le riserve necessarie per lo sviluppo precoce dell’embrione. Durante lo sviluppo, dopo la riorganizzazione dei microtubuli e il trasporto asimmetrico dei determinanti materni, l’ovocita si polarizza lungo gli assi antero-dorsale e dorso-ventrale. Questi assi determinano i successivi assi di polarità dell’embrione e dell’adulto derivanti dalla fecondazione di questo ovocita1. Durante l’oogenesi, il nucleo adotta una posizione asimmetrica nell’ovocita. Nella fase 6, il nucleo è centrato nella cellula. Su un segnale ancora da identificare emesso dalle cellule follicolari posteriori che viene ricevuto dall’ovocita, il nucleo migra verso l’intersezione tra le membrane plasmatiche anteriore e laterale nello stadio 7 (Figura 1B)2,3. Questa posizione asimmetrica è necessaria per indurre la determinazione dell’asse dorso-ventrale.
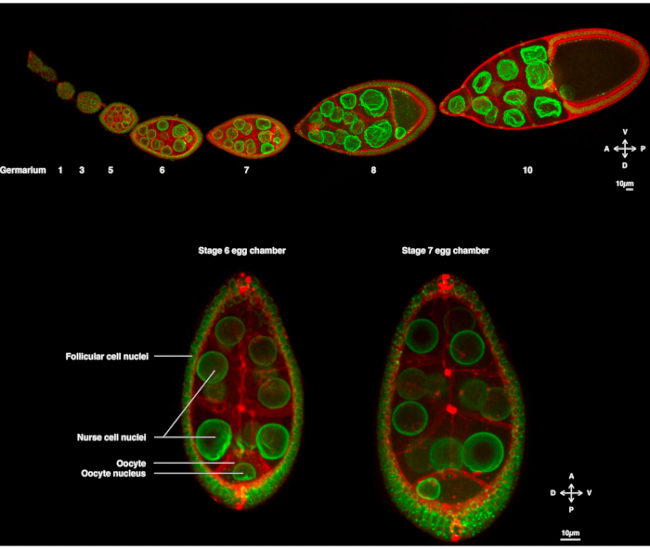
Figura 1: Drosophila melanogaster camere d’uovo. (A) Ovariole fisso da mosche transgeniche che esprimono Fs(2)Ket-GFP che etichetta gli involucri nucleari e ubi-PH-RFP che etichetta le membrane plasmatiche. L’ovariole è composto da camere per uova in via di sviluppo in diverse fasi. La maturazione aumenta lungo l’asse antero-posteriore con il germario sulla punta anteriore (a sinistra) dove risiede la cellula staminale germinale e lo stadio più vecchio sulla punta posteriore (a destra). (B) Proiezione Z della camera dell’uovo vivente mediante microscopia confocale a disco rotante allo stadio 6 dell’oogenesi (a sinistra), in cui il nucleo è centrato nell’ovocita. Il nucleo migrerà per adottare una posizione asimmetrica allo stadio 7 (a destra) a contatto con la membrana plasmatica anteriore (tra l’ovocita e la cellula nutrice) e la membrana plasmatica laterale (tra l’ovocita e le cellule follicolari). Questa posizione indurrà la determinazione del lato dorsale e, quindi, dell’asse dorso-ventrale della camera dell’uovo. Fare clic qui per visualizzare una versione più grande di questa figura.
Per molti decenni, questa migrazione nucleare è stata studiata su tessuti fissi mediante immuno colorazione. Questo approccio ha notevolmente permesso di dimostrare che questo processo dipende da una fitta rete di microtubuli4,5. Più recentemente, abbiamo sviluppato un protocollo che offre condizioni compatibili con l’imaging dal vivo dell’ovocita per diverse ore che consentono di studiare questo processo in modo dinamico6.
Quindi, per la prima volta, siamo stati in grado di descrivere che il nucleo ha traiettorie preferenziali e caratteristiche durante la sua migrazione, una lungo la membrana plasmatica anteriore (APM) e un’altra lungo la membrana plasmatica laterale (LPM) dell’ovocita (Figura 2). Questi ultimi risultati sottolineano l’importanza dei protocolli di live-imaging quando si studiano processi dinamici come la migrazione nucleare.
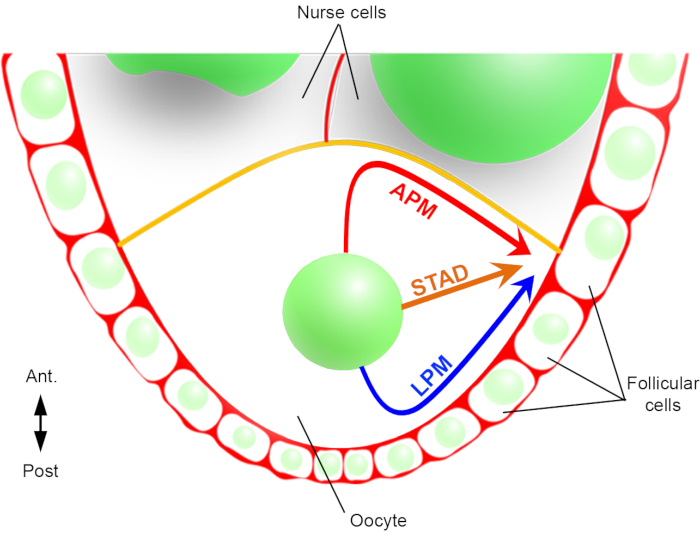
Figura 2: Rappresentazione schematica dei diversi percorsi di migrazione del nucleo. Allo stadio 6 dell’oogenesi, l’ovocita è una grande cellula con un nucleo centrale. In questa fase, l’asse di polarità antero-posteriore è impostato con una membrana plasmatica posteriore/laterale dell’ovocita a contatto con le cellule follicolari e la membrana plasmatica anteriore (in giallo) è in contatto con le cellule nutrici2. Abbiamo precedentemente riportato che il nucleo potrebbe migrare lungo la membrana plasmatica anteriore (APM), lungo la membrana plasmatica laterale (LPM), o attraverso il citoplasma (STAD, direttamente alla corteccia antero-dorsale)6. Fare clic qui per visualizzare una versione più grande di questa figura.
La migrazione del nucleo ovocitario è un fenomeno di circa 3 h6e finora l’evento che innesca l’inizio della migrazione effettiva è sconosciuto. L’inizio della migrazione può anche essere ritardato dai mutanti proteici utilizzati per studiare questo meccanismo. Queste variabili sconosciute ci hanno motivato ad acquisire immagini per lunghi periodi di tempo (10-12 ore). È quindi importante garantire che gli ovociti rimangano vivi. Man mano che la camera dell’uovo si sviluppa, si allunga lungo l’asse antero-posteriore da una forma sferica a una ellittica. Questo allungamento è guidato dalla rotazione delle cellule follicolari, che si verifica dallo stadio 1 allo stadio 8, perpendicolare all’asse antero-posteriore7. Inoltre, una guaina tubolare di muscolo con proprietà pulsatile circonda le camere delle uova. La sua funzione fisiologica è quella di spingere i follicoli in via di sviluppo verso l’ovidotto continuamente8. Al fine di limitare i movimenti che inducono oscillazioni delle camere delle uova dopo la loro dissezione, abbiamo progettato una microcamera di osservazione di 150 μm di altezza (Figura 3A). Questa altezza è leggermente superiore alla dimensione di un follicolo negli stadi 10 e 11. Limita considerevolmente i movimenti verticali del campione preservando la rotazione della camera delle uova, con conseguenti difetti limitati nello sviluppo del follicolo. Eseguiamo quindi l’imaging dal vivo per 12 ore su camere di uova sezionate mediante acquisizioni time-lapse multi-posizione utilizzando un microscopio confocale a disco rotante. Qui descriviamo il nostro protocollo per studiare la migrazione nucleare degli ovociti tra gli stadi 6 e 7.
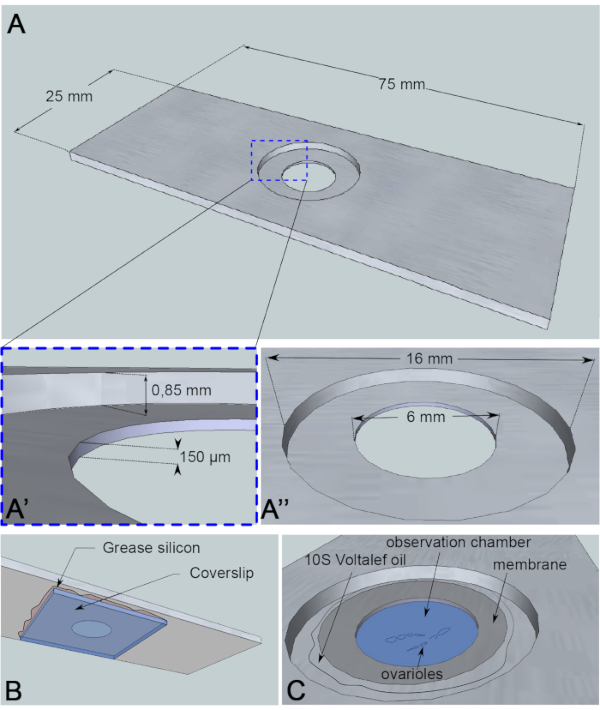
Figura 3: Rappresentazione schematica della camera di osservazione. (A) (Vista dall’alto) Dimensioni precise della slitta di alluminio con le altezze (A’) e le circonferenze (A”) del pozzo perforato al centro della slitta. (B) (vista in basso) Un coperchio che blocca il pozzo è sigillato al vetrino con grasso di silicio. (C) (Vista dall’alto) Gli ovarioli sezionati si sviluppano in un mezzo di imaging coperto da una membrana permeabile ai gas. L’olio di alocarbonio viene utilizzato per stabilizzare la membrana. Fare clic qui per visualizzare una versione più grande di questa figura.
Per seguire la migrazione nucleare e valutare con precisione le traiettorie nell’ovocita, sono necessari marcatori sia per l’involucro nucleare che per la membrana plasmatica. A tal fine, sono stati selezionati due transgeni che hanno un elevato rapporto segnale/rumore e non svaniscono nel corso dell’imaging dal vivo. Per etichettare la membrana plasmatica, si raccomanda l’uso di un P[ubi-PH-RFP] che codifica il dominio Pleckstrin Homology (PH) della Fosfolipasi C Umana ∂1 (PLC∂1) fuso a RFP. Questo dominio PH si lega al fosfoinositide PI(4,5)P2 distribuito lungo la membrana plasmatica dell’ovocita9. Per l’involucro nucleare, il ceppo di trappola proteica P[PPT-un1]Fs(2)Ket-GFP in cui GFP è inserito all’interno del gene che codifica per la Drosophila ß-importina mostra un segnale omogeneo e intenso10. Le giovani mosche (1-2 giorni) vengono poste in fiale fresche contenenti lievito secco 24-48 ore prima della dissezione ovarica.
Per questo test di live-imaging, un pezzo di alluminio di 1 mm di spessore, che non è reattivo per il campione, è stato tagliato nelle dimensioni di un vetrino di microscopia. Ha un foro di 16 mm di diametro al centro della slitta che è stato controforato a 0,85 mm. Questo foro di controforo ha un foro aggiuntivo di 6 mm di diametro con una profondità di 150 μm (Figura 3A). Un coverslip è incollato con grasso siliconico (inerte per il campione) nella parte inferiore della camera di alluminio (Figura 3B). Dopo aver posizionato i campioni nel pozzo riempito a medio, una membrana permeabile allo scambio O2/ CO2 viene posizionata sul mezzo e circondata da olio di alocarbonio (Figura 3C).
Per la dissezione, si consiglia di utilizzare pinci in acciaio inossidabile con una dimensione della punta di 0,05 x 0,02 mm e aghi di diametro 0,20 mm per la separazione degli ovarioli (Figura 4B,C). I nuclei migratori sono ripresi su un microscopio confocale invertito a disco rotante CSU-X1 dotato di una telecamera. Le immagini multi-posizione sono state acquisite mediante time-lapse ogni 15 minuti a 24 °C. Un intervallo di 15 minuti consente di eseguire acquisizioni multi-posizione con fotosbiancamento limitato delle proteine fluorescenti e fototossicità per i campioni. Inoltre, un intervallo più breve non fornirebbe dati molto più informativi per seguire le traiettorie nucleari. I filmati vengono elaborati e analizzati tramite il software Fiji11.
