Durante varios años, el ovocito Drosophila ha surgido como un sistema modelo para estudiar la migración nuclear. El ovocito Drosophila se desarrolla en una estructura multicelular llamada cámara de óvulos. Las cámaras de óvulos abarcan 16 células germinales (15 células nodriza y el ovocito) rodeadas por una capa epitelial de células somáticas foliculares. El desarrollo de la cámara de óvulos se ha subdividido en 14 etapas(Figura 1A),durante las cuales el ovocito crecerá y acumulará las reservas necesarias para el desarrollo temprano del embrión. Durante el desarrollo, tras la reorganización de los microtúbulos y el transporte asimétrico de los determinantes maternos, el ovocito se polariza a lo largo de los ejes antero-dorsal y dorso-ventral. Estos ejes determinan los ejes de polaridad posteriores del embrión y del adulto derivados de la fecundación de este ovocito1. Durante la oogénesis, el núcleo adopta una posición asimétrica en el ovocito. En la etapa 6, el núcleo está centrado en la célula. Tras una señal aún no identificada emitida por las células foliculares posteriores que es recibida por el ovocito, el núcleo migra hacia la intersección entre las membranas plasmáticas anterior y lateral en la etapa 7(Figura 1B)2,3. Esta posición asimétrica es necesaria para inducir la determinación del eje dorso-ventral.
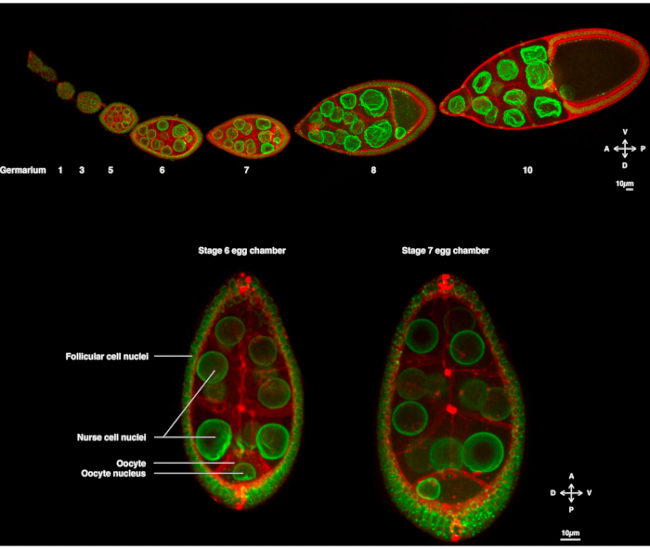
Figura 1: Cámaras de huevo de Drosophila melanogaster. (A) Ovariole fijo de moscas transgénicas que expresan Fs(2)Ket-GFP que etiqueta las envolturas nucleares y ubi-PH-RFP que marca las membranas plasmáticas. El ovariole se compone de cámaras de huevo en desarrollo en diferentes etapas. La maduración aumenta a lo largo del eje antero-posterior con el germario en la punta anterior (izquierda) donde reside la célula madre germinal y la etapa más antigua en la punta posterior (derecha). (B) Proyección Z de la cámara de huevo vivo mediante microscopía confocal de disco giratorio en la etapa 6 de la oogénesis (izquierda), en la que el núcleo se centra en el ovocito. El núcleo migrará para adoptar una posición asimétrica en la etapa 7 (derecha) en contacto con la membrana plasmática anterior (entre el ovocito y la célula nodriza) y la membrana plasmática lateral (entre el ovocito y las células foliculares). Esta posición inducirá la determinación del lado dorsal y, por lo tanto, del eje dorso-ventral de la cámara de huevos. Haga clic aquí para ver una versión más grande de esta figura.
Durante muchas décadas, esta migración nuclear se ha estudiado en tejidos fijos mediante inmunotinción. Este enfoque ha hecho notablemente posible demostrar que este proceso depende de una densa red de microtúbulos4,5. Más recientemente, desarrollamos un protocolo que ofrece condiciones compatibles con la obtención de imágenes en vivo del ovocito durante varias horas, lo que permite estudiar este proceso dinámicamente6.
De ahí que, por primera vez, hayamos podido describir que el núcleo tiene trayectorias preferentes y características durante su migración, una a lo largo de la membrana plasmática anterior (APM) y otra a lo largo de la membrana plasmática lateral (LPM) del ovocito(Figura 2). Estos últimos resultados subrayan la importancia de los protocolos de imágenes en vivo cuando se estudian procesos dinámicos como la migración nuclear.
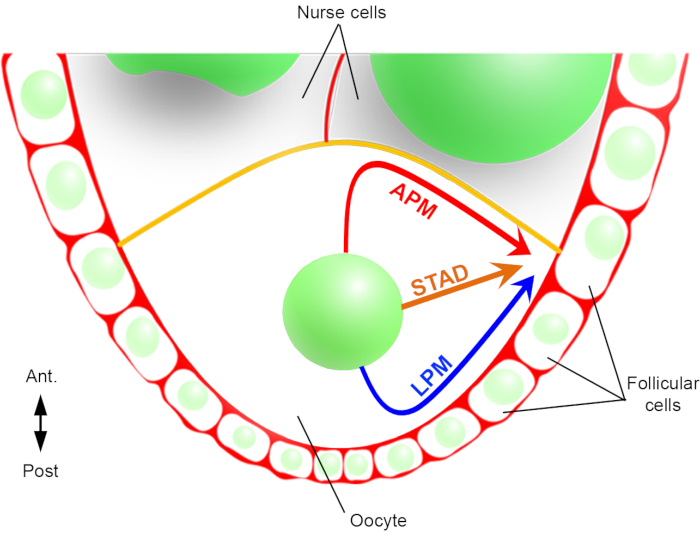
Figura 2: Representación esquemática de las diferentes rutas de migración del núcleo. En la etapa 6 de la oogénesis, el ovocito es una célula grande con un núcleo central. En esta etapa, el eje de polaridad antero-posterior se establece con una membrana plasmática posterior/lateral del ovocito en contacto con las células foliculares y la membrana plasmática anterior (en amarillo) está en contacto con las células nodriza2. Hemos informado previamente que el núcleo podría migrar a lo largo de la membrana plasmática anterior (APM), a lo largo de la membrana plasmática lateral (LPM), o a través del citoplasma (STAD, directamente a la corteza antero-dorsal)6. Haga clic aquí para ver una versión más grande de esta figura.
La migración del núcleo de ovocitos es un fenómeno de aproximadamente 3 h6, y hasta ahora, se desconoce el evento que desencadena el inicio de la migración real. El inicio de la migración también puede ser retrasado por los mutantes de proteínas utilizados para estudiar este mecanismo. Estas variables desconocidas nos motivaron a adquirir imágenes durante largos períodos de tiempo (10-12 h). Por lo tanto, es importante asegurarse de que los ovocitos permanezcan vivos. A medida que la cámara de huevos se desarrolla, se alarga a lo largo del eje antero-posterior de una forma esférica a una elíptica. Este alargamiento es impulsado por la rotación de las células foliculares, que ocurre desde la etapa 1 hasta la etapa 8, perpendicular al eje antero-posterior7. Además, una hela tubular de músculo con propiedad pulsátil rodea las cámaras de huevos. Su función fisiológica es empujar los folículos en desarrollo hacia el oviducto continuamente8. Con el fin de limitar los movimientos que inducen oscilaciones de las cámaras de huevos después de su disección, diseñamos una microcámara de observación que mide 150 μm de altura(Figura 3A). Esta altura es marginalmente más alta que el tamaño de un folículo en las etapas 10 y 11. Limita considerablemente los movimientos verticales de la muestra al tiempo que preserva la rotación de la cámara de óvulos, lo que resulta en defectos limitados en el desarrollo del folículo. Luego realizamos imágenes en vivo durante 12 h en cámaras de huevos diseccionadas mediante adquisiciones de lapso de tiempo de múltiples posiciones utilizando un microscopio confocal de disco giratorio. Aquí describimos nuestro protocolo para estudiar la migración nuclear de ovocitos entre las etapas 6 y 7.
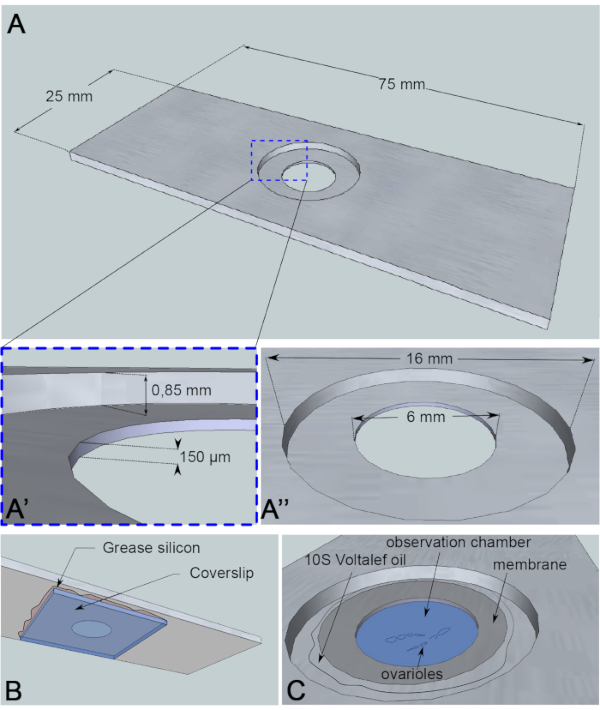
Figura 3: Representación esquemática de la cámara de observación. (A) (Vista superior) Dimensiones precisas de la corredera de aluminio con las alturas (A’) y circunferencias (A”) del pozo perforado en el centro de la diapositiva. (B) (Vista inferior) Un codal que bloquea el pozo se sella a la corredera con grasa de silicio. (C) (Vista superior) Los ovarioles disecados se desarrollan en un medio de imagen que está cubierto por una membrana permeable al gas. El aceite de halocarbono se utiliza para estabilizar la membrana. Haga clic aquí para ver una versión más grande de esta figura.
Para seguir la migración nuclear y evaluar con precisión las trayectorias en el ovocito, se necesitan marcadores tanto para la envoltura nuclear como para la membrana plasmática. Con este objetivo, se han seleccionado dos transgenes que tienen una alta relación señal/ruido y no se desvanecen en el transcurso de las imágenes en vivo. Para etiquetar la membrana plasmática, se recomienda el uso de una P[ubi-PH-RFP] que codifique el dominio de Homología de Pleckstrin (PH) de la Fosfolipasa C Humana ∂1 (PLC∂1) fusionada a RFP. Este dominio PH se une al fosfoinosítido PI(4,5)P2 distribuido a lo largo de la membrana plasmática del ovocito9. Para la envoltura nuclear, la cepa trampa de proteína P[PPT-un1]Fs(2)Ket-GFP donde GFP se inserta dentro del gen que codifica la ß-importina de Drosophila muestra una señal homogénea e intensa10. Las moscas jóvenes (1-2 días de edad) se colocan en viales frescos que contienen levadura seca 24-48 h antes de la disección del ovario.
Para este ensayo de imágenes en vivo, se ha cortado una pieza de aluminio de 1 mm de espesor, que no es reactiva para la muestra, en las dimensiones de una diapositiva de microscopía. Tiene un orificio de 16 mm de diámetro en el centro de la corredera que ha sido contraborrado a 0,85 mm. Este contrabordo tiene un orificio adicional de 6 mm de diámetro con una profundidad de 150 μm (Figura 3A). Un coverslip se pega con grasa de silicona (inerte para la muestra) en la parte inferior de la cámara de aluminio(Figura 3B). Después de colocar las muestras en el pozo lleno de medio, se coloca una membrana permeable al intercambio de O2/ CO2 sobre el medio y rodeada de aceite de halocarbono(Figura 3C).
Para la disección, se recomienda utilizar fórceps de acero inoxidable con una dimensión de punta de 0.05 x 0.02 mm, y agujas de 0.20 mm de diámetro para la separación de los ovarioles(Figura 4B,C). Los núcleos migratorios se capturan en un microscopio invertido confocal de disco giratorio CSU-X1 equipado con una cámara. Las imágenes multi-posición fueron adquiridas por time-lapse cada 15 min a 24 °C. Un intervalo de 15 minutos permite realizar adquisiciones multi-posición con fotobleaching limitado de las proteínas fluorescentes y fototoxicidad para las muestras. Además, un intervalo más corto no proporcionaría datos mucho más informativos para seguir las trayectorias nucleares. Las películas se procesan y analizan a través del software Fiji11.
