I flera år har Drosophila-äggoiden dykt upp som ett modellsystem för att studera nukleär migration. Drosophila oocyte utvecklas i en multicellulär struktur som kallas äggkammaren. Äggkammare omfattar 16 könsceller (15 sjuksköterskeceller och oocyt) omgivna av ett epiteliallager av follikulära somatiska celler. Äggkammarutvecklingen har delats in i 14 steg (figur 1A), under vilken äggcellen kommer att växa och ackumulera reserver som är nödvändiga för embryots tidiga utveckling. Under utvecklingen, vid mikrotubule omorganisering och asymmetrisk transport av moderns bestämningsfaktorer, polariseras oocyten längs antero-dorsala och dorso-ventrala axlar. Dessa axlar bestämmer embryots och den vuxnas efterföljande polaritetsaxlar som härrör från befruktningen av denna oocyt1. Under oogenes antar kärnan en asymmetrisk position i äggcellen. I steg 6 är kärnan centrerad i cellen. Vid en ännu inte identifierad signal som avges av de bakre follikulära cellerna och som tas emot av äggcellen migrerar kärnan mot skärningspunkten mellan de främre och laterala plasmamembranen i steg 7 (figur 1B)2,3. Denna asymmetriska position krävs för att inducera bestämningen av dorso-ventralaxeln.
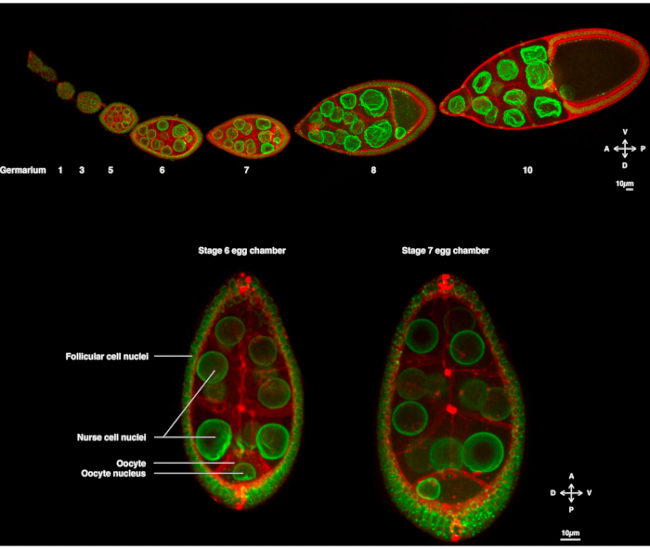
Figur 1: Drosophila melanogaster äggkammare. (A) Fast ovariole från transgena flugor som uttrycker Fs(2)Ket-GFP som märker de nukleära kuvert och ubi-PH-RFP som märker plasmamembranen. Ovariolen består av att utveckla äggkammare i olika stadier. Mognad ökar längs den antero-bakre axeln med pelargonen vid den främre spetsen (vänster) där bakteriestamcellen bor och det äldre stadiet vid den bakre spetsen (höger). B)Z-projektion av levande äggkammare genom att snurra diskkonfokal mikroskopi i steg 6 av oogenes (vänster), där kärnan är centrerad i äggcellen. Kärnan migrerar för att anta en asymmetrisk position i steg 7 (höger) i kontakt med det främre plasmamembranet (mellan äggcellen och sjuksköterskans cell) och det laterala plasmamembranet (mellan äggcellen och follikulära cellerna). Denna position kommer att inducera bestämningen av dorsala sidan och därmed äggkammarens dorso-ventrala axel. Klicka här om du vill visa en större version av den här figuren.
I många årtionden har denna nukleära migration studerats på fasta vävnader genom immunostaining. Detta tillvägagångssätt har särskilt gjort det möjligt att visa att denna process beror på ett tätt nätverk av mikrotubuli4,5. Mer nyligen utvecklade vi ett protokoll som erbjuder villkor som är kompatibla med live imaging av äggcellen under flera timmar vilket gör det möjligt att studera denna process dynamiskt6.
Därför har vi för första gången kunnat beskriva att kärnan har förmånliga och karakteristiska banor under sin migration, en längs det främre plasmamembranet (APM) och en annan längs äggcellens laterala plasmamembran (LPM) i äggcellen (figur 2). Dessa senaste resultat understryker vikten av live-imaging protokoll när man studerar dynamiska processer såsom kärnmigration.
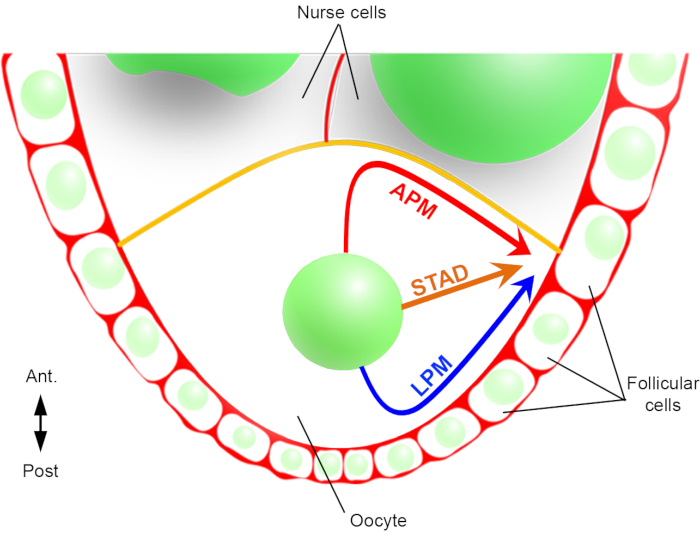
Figur 2: Schematisk representation av kärnans olika migreringsbanor. I steg 6 av oogenesen är oocyten en stor cell med en central kärna. I detta skede ställs den antero-bakre polaritetsaxeln in med ett bakre/laterala plasmamembran av oocytet i kontakt med follikulära celler och det främre plasmamembranet (i gult) är i kontakt med sjuksköterskans celler2. Vi har tidigare rapporterat att kärnan kan migrera antingen längs det främre plasmamembranet (APM), längs det laterala plasmamembranet (LPM), eller genom cytoplasman (STAD, direkt till antero-dorsala cortex)6. Klicka här om du vill visa en större version av den här figuren.
Oocytkärnan migration är ett fenomen på ca 3 h6, och hittills är händelsen som utlöser början av den faktiska migreringen okänd. Starten av migreringen kan också försenas av proteinmutanter som används för att studera denna mekanism. Dessa okända variabler motiverade oss att förvärva bilder under långa tidsperioder (10-12 h). Det är därför viktigt att se till att äggcellerna förblir vid liv. När äggkammaren utvecklas sträcker den sig längs den antero-bakre axeln från en sfärisk till en elliptisk form. Denna förlängning drivs av rotationen av follikulära celler, som uppstår från steg 1 till steg 8, vinkelrätt mot antero-bakre axeln7. Dessutom omger en rörformig muskelstel med pulsatil egenskap äggkamrarna. Dess fysiologiska funktion är att driva de utvecklande folliklarna mot ovidukten kontinuerligt8. För att begränsa de rörelser som inducerar svängningar i äggkamrarna efter deras dissekering konstruerade vi en observationsmikrokammare som mäter 150 μm i höjd (figur 3A). Denna höjd är marginellt högre än storleken på en follikel i steg 10 och 11. Det begränsar avsevärt provets vertikala rörelser samtidigt som äggkammarens rotation bevaras, vilket resulterar i begränsade defekter i follikelutvecklingen. Vi utför sedan live imaging i 12 h på dissekerade äggkammare genom flerpositions timelapse förvärv med hjälp av en spinning-disk confocal mikroskop. Här beskriver vi vårt protokoll för att studera oocyt nukleär migration mellan steg 6 och 7.
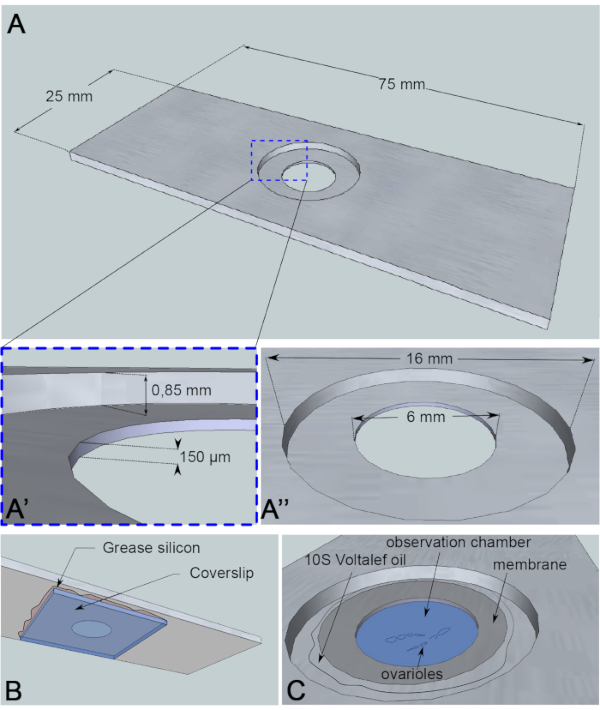
Figur 3:Schematisk representation av observationskammaren. (A) (Överst) Exakta dimensioner på aluminiumrutschbanan med höjderna (A’) och omkretsarna (A”) på brunnen som borras i mitten av bilden. (B) (Nederkant) Ett täckglas som blockerar brunnen förseglas till diabilden med silikonfett. (C) (Top view) Dissekerade ovarioler utvecklas i ett bildmedium som täcks av ett gaspermeabelt membran. Halokarbonolja används för att stabilisera membranet. Klicka här om du vill visa en större version av den här figuren.
För att följa nukleär migration och exakt bedöma banor i äggcellen behövs markörer för både kärnhöljet och plasmamembranet. Med detta syfte har två transgener som har ett högt signal-/brusförhållande och inte bleknar under loppet av live-avbildning valts. För att märka plasmamembranet rekommenderas användning av en P[ubi-PH-RFP] som kodar pleckstrin Homology (PH) domänen för humanfosfolipas C ∂1 (PLC∂1) som smälts till RFP. Denna PH-domän binder till fosfoinositide PI(4,5)P2 fördelad längs plasmamembranet i äggcellen9. För kärnhöljet visar P[PPT-un1]Fs(2)Ket-GFP proteinfälla stam där GFP sätts in i genen kodning Drosophila ß-importin en homogen och intensiv signal10. Unga flugor (1-2 dagar gamla) placeras i färska injektionsflaskar som innehåller torr jäst 24-48 h före äggstocksdesektion.
För denna levande avbildningsanalys har en 1 mm tjock bit aluminium, som inte är reaktiv för provet, skurits i dimensionerna av en mikroskopibild. Den har ett hål med diametern 16 mm i mitten av diabilden som har kontraborerats till 0,85 mm. Denna räknare har ytterligare ett hål med diametern 6 mm med ett djup på 150 μm (figur 3A). Ett täckglas limmas med silikonfett (inert för provet) längst ner i aluminiumkammaren (Figur 3B). Efter att proverna placerats i den medelfyllda brunnen placeras ettmembransom är genomsläppligt till O 2 /CO2-utbytet över mediet och omges av halokarbonolja (figur 3C).
För dissekeringen rekommenderas att använda rostfria tångar med en spetsdimension på 0,05 x 0,02 mm och nålar med diametern 0,20 mm för separation av ovariolerna(figur 4B,C). De migrerande kärnorna avbildas på ett konvertiskt inverterat mikroskop med spinnskiva som är utrustat med en kamera. Flerpositionsbilder förvärvades genom timelapse var 15: e minut vid 24 °C. Ett intervall på 15 minuter gör det möjligt att utföra flerpositionsförvärv med begränsad fotoblekning av fluorescerande proteiner och fototoxicitet för proverna. Dessutom skulle ett kortare intervall inte ge mycket mer informativa uppgifter för att följa kärnkraftsbanorna. Filmerna bearbetas och analyseras via Fiji programvara11.
