Studies were carried out on male and female c57Bl/6 mice aged 3-12 months. All experimental procedures were performed in accordance with the University of Newcastle's Animal Care and Ethics Committee (protocols A-2013-312, and A-2020-002).
1. In vitro electrophysiology
- Preparation of solutions for spinal cord slice preparation and recording
- Artificial cerebrospinal fluid
NOTE: Artificial cerebrospinal fluid (aCSF) is used in an interface incubation chamber, where slices are stored until recording commences and during experiments as both perfusate and diluent for drugs. See Table 1 for the detailed composition.
- Artificial cerebrospinal fluid
| Chemical | aCSF (mM) | aCSF (g/100 mL) | Sucrose-substituted aCSF (mM) | Sucrose-substituted aCSF (g/100 mL) | High-potassium aCSF (mM) | High-potassium aCSF (g/100 mL) |
| Sodium chloride (NaCl) | 118 | 0.690 | – | – | 118 | 0.690 |
| Sodium hydrogen carbonate (NaHCO3) | 25 | 0.210 | 25 | 0.210 | 25 | 0.210 |
| Glucose | 10 | 0.180 | 10 | 0.180 | 10 | 0.180 |
| Potasium chloride (KCl) | 2.5 | 0.019 | 2.5 | 0.019 | 4.5 | 0.034 |
| Sodium dihydrogen phosphate (NaH2PO4) | 1 | 0.012 | 1 | 0.012 | 1 | 0.012 |
| Magnesium cloride (MgCl2) | 1 | 0.01 | 1 | 0.01 | 1 | 0.01 |
| Calcium chloride (CaCl2) | 2.5 | 0.028 | 2.5 | 0.028 | 2.5 | 0.028 |
| Sucrose | – | – | 250 | 8.558 | – | – |
Table 1: Artificial Cerebrospinal Fluid compositions. Abbreviation: aCSF = artificial cerebrospinal fluid.
- Prepare aCSF containing (in mM) 118 NaCl, 25 NaHCO3, 10 glucose, 2.5 KCl, 1 NaH2PO4, 1 MgCl2, and 2.5 CaCl2 by adding the required quantities of the above, excluding CaCl2, to 2 L of distilled water.
- Bubble the above solution with carbogen (95% O2, 5% CO2) for 5 min and add CaCl2.
NOTE: This step prevents CaCl2 precipitation, i.e., the solution should not turn cloudy. For drug application during experiments, dilute the drug stock solutions in aCSF to desired final concentrations.
- Sucrose-substituted artificial cerebrospinal fluid
NOTE: Sucrose-substituted aCSF is used during dissection and spinal cord slicing. As indicated by the name, sucrose is substituted for NaCl to reduce neuronal excitation during these procedures while maintaining osmolarity. See Table 1 for the detailed composition.- Prepare sucrose-substituted aCSF containing (in mM) 250 sucrose, 25 NaHCO3, 10 glucose, 2.5 KCl, 1 NaH2PO4, 1 MgCl2, and 2.5 CaCl2 by adding the required quantities of all of the above, excluding CaCl2, to 300 mL of distilled water.
- Bubble the solution with carbogen for 5 min and then add CaCl2.
- Store the solution in a -80 °C freezer for approximately 40 min or until the solution forms a slurry. Avoid freezing solid and use while in slurry consistency.
- Microelectrode array preparation
NOTE: The contact surface of the MEA requires a pretreatment to make it hydrophilic.- Before the experiment, fill the MEA well with either fetal bovine serum (FBS) or horse serum (HS) for 30 min.
- Remove the FBS or HS and thoroughly rinse MEA with approximately five washes of distilled water until the distilled water is no longer foamy. Fill the well with aCSF, ready for use.
- Acute spinal cord slice preparation
NOTE: The mouse spinal cord slice preparation is as previously described by Smith et al.2. Ideally, removal of the lumbosacral enlargement should take no more than 8-10 min (steps 1.3.2-1.3.11 below).- Deeply anesthetize the mouse with 100 mg/kg ketamine (i.p.) and then decapitate it using large surgical scissors.
- Remove the skin over the abdominal region by making a small cut in the skin at the level of the hips. Pull the skin on either side of the cut rostrally until all the skin is removed, i.e., from the top of the rib cage to the top of the pelvis (both ventrally and dorsally).
- Place the body on ice and use a ventral approach to expose the vertebral column by removing all the viscera and cutting through the ribs lateral to the sternum.
- Remove the ventral rib cage, both scapulae (cut off at approximately T2), and the lower limbs and pelvis (cut off at approximately the top of the sacrum).
- Transfer the vertebral column and rib preparation to a dissecting bath containing ice-cold sucrose aCSF. Pin all four corners of the preparation (ventral surface upwards) by placing pins through the lower back muscles and the attached upper ribs.
- Remove all muscle and connective tissue overlying the ventral surface of the vertebrae with rongeurs and identify the vertebral region over the lumbosacral enlargement, which lies approximately beneath the T12 to L2 vertebral bodies.
- Remove a vertebral body that is caudal to the lumbosacral enlargement region to provide access to the spinal cord as it sits in the vertebral canal.
- Using curved spring scissors, cut through the vertebral pedicles bilaterally while lifting and pulling the vertebral body rostrally to separate the ventral and dorsal aspects of the vertebrae and expose the spinal cord.
- Once the vertebral bodies are removed to reveal the lumbosacral enlargement, carefully clear the remaining roots that anchor the spinal cord with spring scissors until the cord floats free.
- Isolate the spinal cord with rostral and caudal cuts well above and below the lumbosacral enlargement, allowing the target region of the cord to 'float free.'
NOTE: The preferred slice orientation will determine how the cord is subsequently mounted for sectioning (Figure 1). - For transverse slices, lift the lumbosacral segment by an attached root and place it on a pre-cut polystyrene (Styrofoam) block (1 cm x 1 cm x 1 cm) with a shallow channel cut in the center. Use cyanoacrylate adhesive (see the Table of Materials) to attach the block and cord to the sectioning platform and place it in the cutting bath containing ice-cold sucrose aCSF (slurry).
NOTE: The shallow channel helps secure and orient the spinal cord, with the dorsal side exposed and the thoracic end of the cord at the bottom of the block. - For sagittal slices, lay a thin line of cyanoacrylate adhesive on the sectioning platform, lift the lumbosacral enlargement by an attached root, and place the cord along the line of glue, ensuring one lateral surface is in the adhesive and the other faces upwards. Place it in the cutting bath containing ice-cold sucrose aCSF (slurry).
- For horizontal slices, put a thin line of cyanoacrylate adhesive on the sectioning platform. Lift the lumbosacral enlargement by an attached root, and place the lumbosacral enlargement along the line of adhesive, ensuring the ventral surface is in the adhesive and the dorsal surface faces upwards. Use attached roots to position the cord. Place it in the cutting bath containing ice-cold sucrose aCSF (slurry).
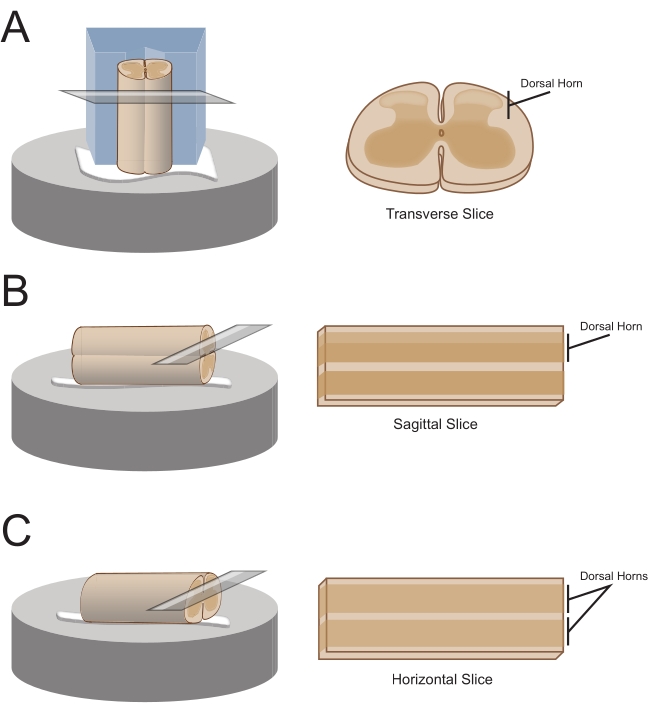
Figure 1: Spinal cord slice orientations, mounting and cutting methods. (A) Transverse slices require a Styrofoam cutting block with a supporting groove cut into it. The spinal cord is rested against the block in the support groove, the dorsal side of the cord facing away from the block. The block and cord are glued onto a cutting stage with cyanoacrylate adhesive. (B) Sagittal slices are prepared by placing a thin line of cyanoacrylate adhesive on the cutting stage and then positioning the spinal cord on its side on the glue. (C) Horizontal slices are prepared by placing a thin line of cyanoacrylate adhesive on the cutting stage and then positioning the spinal cord ventral side down on the glue. Please click here to view a larger version of this figure.
- Obtain 300 µm thick slices (L1-L5, same thickness regardless of orientation) using a vibrating microtome with the following settings: speed 0.06 mm/s, amplitude 2.50 mm, and calibrated to within ±0.02 height amplitude deviation.
- Transfer the slices to an air interface incubation chamber containing oxygenated aCSF.
- Before recording, allow the slices to equilibrate for 1 h at room temperature (20-24 °C).
- Microelectrode array recordings
NOTE: The following steps detail how to use record data from MEA-based experiments on spinal cord slices. Several MEA designs can be used depending on the experiment. Design details for MEAs used in these experiments are shown in Table 2 and Figure 2. Detailed design information has been published by Egert et al.26 and Thiebaud et al.27 for planar and 3-dimensional (3D) MEAs, respectively. Both MEA types are composed of 60 titanium nitride electrodes, with a silicon nitride insulating layer and titanium nitride tracks and contact pads.- Experimental setup
- Turn on the computer and interface board, and start the recording software.
- Load the pre-assembled recording template (Figure 3A). Name the files for the day in the recorder tab.
- Continuously bubble aCSF with carbogen (5% CO2, 95% O2) for the duration of the experiment.
- Turn the perfusion system on, which is controlled by a peristaltic pump. Place the inlet line into aCSF and the inlet end in a waste beaker. Prime the perfusion lines with aCSF.
- Prepare 4-AP and any other drug solutions by diluting stocks in 50 mL of aCSF to the required final concentration (e.g., 200 µM for 4-AP).
- Place the drug solutions in drug pots and bubble them with carbogen.
- 4-AP activity
- Following incubation, transfer a single slice from the incubator using a large-tip Pasteur pipette filled with aCSF.
- Place the slice in the MEA well and add additional aCSF.
- Position the slice over the 60-electrode recording array using a fine short hair paintbrush. Avoid contacting the electrodes with the paintbrush or dragging the tissue across the electrodes, especially if using 3D arrays.
NOTE: Depending on the MEA layout, this can be done with or without the assistance of a microscope for accurate positioning. - After positioning the slice, place a weighted net over the tissue to hold it in place and promote good contact with MEA electrodes.
NOTE: The slice may need repositioning following net placement. - Place the MEA in the recording headstage (Figure 2A,B).
- Check the position of the tissue over the electrodes using an inverted microscope (2x magnification) to confirm that as many electrodes as possible are under the superficial DH (SDH). Ensure that at least 2-6 electrodes do not contact the slice as these electrodes are important for subtracting noise and recording artefacts during analysis (Figure 2E).
- Turn on the camera, connect it to the device, and take a reference image of the slice relative to the MEA for use during analysis.
- Press Start DAQ in the recording software, and confirm that all electrodes are receiving a clear signal.
NOTE: If the signal is noisy, unclip the headstage, and clean both the MEA contact pads and gold spring contacts with 70% ethanol (use a laboratory wipe to ensure that the pads and contacts are dry after cleaning). If the signal is still noisy, turn off the malfunctioning electrodes in the recording software or note down for exclusion later during analysis. - Attach the perfusion inlet and outlet lines to the MEA-well (previously filled with aCSF) and turn the perfusion system on. Check the flow rate, ideally 4-6 bath volumes per minute, and ensure that the outflow is sufficient to prevent overflow of the superfusate.
- Allow the tissue to equilibrate for 5 min and then record 5 min of raw, unfiltered baseline data.
- Move the perfusion inlet line from aCSF to a 4-AP solution and wait for 12 min for the 4-AP-induced rhythmic activity to reach steady state (2 min for drugs to reach the bath and 10 min for the activity to peak and then plateau).
- Record 5 min of 4-AP-induced activity. Be prepared for subsequent recordings to test the drugs or to check the stability of 4-AP.
- Experimental setup
| Microelectrode Array Layouts | ||||
| Microelectrode Array Model | 60MEA 200/30iR-Ti | 60-3DMEA 100/12/40iR-Ti | 60-3DMEA 200/12/50iR-Ti | 60MEA 500/30iR-Ti |
| Planar or 3-Dimensional (3D) | Planar | 3D | 3D | Planar |
| Electrode Grid | 8 x 8 | 8 x 8 | 8 x 8 | 6 x 10 |
| Electrode Spacing | 200 µm | 100 µm | 200 µm | 500 µm |
| Electrode Diameter | 30 µm | 12 µm | 12 µm | 30 µm |
| Electrode Height (3D) | N/A | 40 µm | 50 µm | N/A |
| Experiments | Transverse slice | Transverse slice | Sagittal + Horizontal | Sagittal + Horizontal |
Table 2: Microelectrode array layouts.
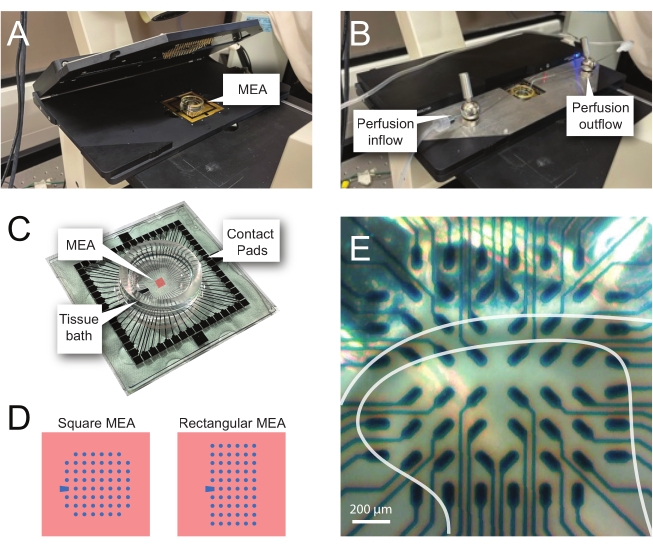
Figure 2: Tissue positioning on the microelectrode array. (A) Image shows an open MEA headstage with an MEA placed in position. (B) Same as A with MEA headstage closed for recordings and tissue perfusion system in place. (C) Image shows an MEA as supplied by the manufacturer. Contact pads, which interface with the gold springs of the headstage, and the MEA tissue bath that holds the tissue bathing solution and tissue slice are shown. The area highlighted by the red square in the center is the location of the electrode array. (D) Schematics show the two MEA electrode configurations used in this study, with further details presented in Table 2. The reference electrode is denoted by the blue trapezoid. The left MEA electrode layout shows a 60-electrode square configuration, used most in the presented work-models 60MEA200/30iR-Ti with 30 μm diameter electrodes spaced 200 μm apart, or 200 μm spaced and 100 μm spaced 3-dimensional MEAs (60MEA200/12/50iR-Ti and 60MEA100/12/40iR-Ti) with electrodes 12 μm in diameter and either 50 μm or 40 μm high, respectively. The left MEA electrode layout shows a 6 x 10 electrode rectangular layout-60MEA500/30iR-Ti. (E) High-magnification image of a 60MEA100/12/40iR-Ti square MEA with transverse spinal cord slice positioned for recording. The slice sits on electrode rows 3-8. The top row of electrodes, which do not contact any tissue, serve as reference electrodes. The SDH area appears as a semitransparent band. In this case, the SDH overlies electrodes in rows 4, 5, and 6 and columns 2, 3, 4, 5, and 7 of the MEA. Scale bar = 200 µm. Abbreviations: MEA = microelectrode array; SDH = superficial dorsal horn. Please click here to view a larger version of this figure.
- Changing slices
- Following each recording session, rinse the lines with aCSF.
- Remove the MEA from the headstage.
- Remove the net and the tissue from the MEA well, rinse them well with aCSF, and repeat the above steps with a new slice.
2. Data processing and analysis
NOTE: The following steps detail how to use the analysis software for MEA experiments on spinal cord slices. One of the 60 electrodes serves as an internal reference (marked by a trapezoid in Figure 2 C,D), while between four and twenty-five of the remaining 59 are positioned under the SDH in an adult mouse spinal cord slice. Subsequent analysis detects extracellular action potential (EAP) and local field potential (LFP) waveforms (see Figure 3B for examples) from the raw signal in this region.
- Raw data processing
- Open the analysis software and load the pre-made analysis layout (Figure 3B).
- Open the file of interest and deselect the reference electrode (electrode 15 in 8 x 8 MEA- or electrode E1 in 6 x 10 MEA-configuration) and any electrodes deemed to be excessively noisy.
- Set the time window for analysis (0:00 → 5:00 min).
- Move to the Cross-channel filter tab. Select Complex reference and select the Reference Electrodes based on the image taken and notes made during the experiment (i.e., those electrodes not under tissue). To apply and check this, press Explore before continuing.
- Move to the EAP filter tab and apply a 2nd order high pass Butterworth filter (200 Hz cut off) to remove LFP activity.
- Move to the LFP filter tab and apply a 2nd order band pass Butterworth filter (delta frequencies of 0.5-4 Hz) to remove EAP activity.
- Move to the EAP detector tab and select Auto threshold. Tick Rising and Falling edge boxes and set the Dead time to 0.5 ms.
- Set Positive and Negative thresholds based on the data. Inspect the data by returning to the Raw data analyzer screen, moving the time marker, and then returning to the EAP detector tab and pressing Explore. Repeat until satisfied that the set detection threshold is capturing EAPs without capturing noise/non-physiological activity. Use the reference electrodes to identify noise/non-physiological activity.
NOTE: It is necessary to ensure a minimal number of EAPs are detected in reference electrodes where physiological activity will not be occurring. However, rather slight deviations in baseline might be falsely detected as EAPs. This is while still aiming to maximize the number of real events detected in the active electrodes. - Move to the LFP detector tab, select the Manual threshold, tick Rising and Falling edge boxes, and set the Dead time to 3 ms.
- Repeat step 2.1.8 for one electrode by selecting an electrode with LFP activity. Once satisfied, select Apply to all as thresholds will only be applied to a single electrode when undertaking manual thresholding.
- While examining LFP data in the Detector tab, note the maximum number of threshold crossings for the one LFP waveform and maximum time separation of threshold crossings for the one LFP waveform for use in later analysis.
- Press Start analysis.
- When the analysis is complete, move to the EAP analyzer tab and export the data. Do the same on the LFP analyzer tab.
- Repeat this process for all other files from the same slice.
- Following data export, convert the files to xlsx format so they can be read by the programming script used. Name the files according to the following convention for the provided script to read them: experiment name (e.g., sample data) – slice number (e.g., S1) – recording number (e.g., R1) – activity type (e.g., spikes or SPs, corresponding to EAPs or LFPs, respectively).
NOTE: The EAP analysis described here treats spiking from individual channels as a single population, even though this activity would commonly arise from multiple neurons in close proximity to the recording electrode. If the number of neurons contributing to EAPs in a channel is desired, multispike sorting techniques described elsewhere can be applied to distinguish distinct populations of spikes based on waveform characteristics28.
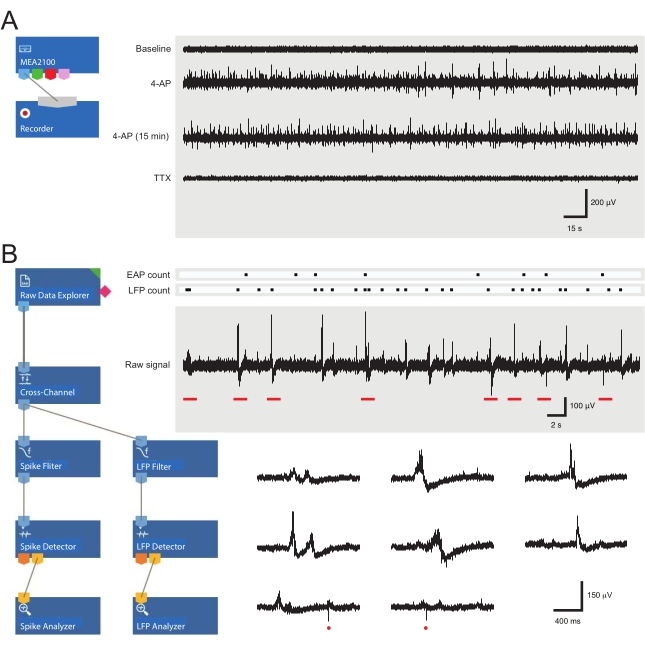
Figure 3: Data recording and analysis tool layouts and example microelectrode array recordings showing extracellular action potential and local field potential waveforms. (A) Schematic shows preconfigured recording templateused for the acquisition of MEA data. Linking the MEA2100 and the recording (headstage/amplifier) tool enables the data to be named and saved. Four example traces of raw data (right, 5-min epochs) were collected by one MEA channel showing activity at baseline, 12 min after 4-AP application, a further 15 min after established 4-AP activity, and following bath application of TTX (1 µM). Note, the addition of 4-AP (second trace) produces a clear increase in background noise and EAP/LFP activity. Importantly, the activity remains relatively stable for at least 15 min after 4-AP-induced activity is established (third trace). Addition of TTX (1 µM) abolishes all activity (bottom trace). (B) Schematic (left) shows analyzer software configuration for data analysis. The raw data explorer tool is used to import recordings collected by recording software. These data are then run through a cross-channel filter tool that subtracts the selected reference electrode(s) signal(s) from other electrodes to remove background noise. Data pass through the EAP filter and the LFP filter tools to optimize signal-to-noise relationships for each waveform. Following this step, the EAP path data enter the EAP detector tool, where thresholds are set. EAPs are detected and then sent to the EAP analyzer tool where the latencies of each event are recorded and exported as a txt. file. An identical workflow occurs for LFP data using a corresponding LFP toolkit. Right traces show data from a single MEA channel containing various extracellular waveforms. Location of EAP and LFP signals are highlighted in the above 'count rasters.' Lower traces are epochs from upper recording (denoted by red bars) showing waveforms on an expanded timescale, including various LFP signals (note the variety of appearances) and individual extracellular EAPs (red circles). Note, LFP/EAP waveform and polarity vary relative to the number of neurons producing these signals, their proximity to the recording electrode, and their location in relation to the nearby electrode(s). Abbreviations: MEA = microelectrode array; EAP = extracellular action potential; LFP = local field potential; 4-AP = 4-aminopyridine; TTX = tetrodotoxin. Please click here to view a larger version of this figure.
- Synchronicity analysis
NOTE: Synchronicity, or the number of 'coincident' events between two electrodes, was determined using the coincidence criterion within the A-SPIKE-synchronization method outlined by Satuvuori et al.29. The script used here only compares electrodes adjacent to one another for efficiency (i.e., horizontal, vertical, and diagonal neighbors); however, the script could be rewritten to compare all electrodes if required.- Perform data analysis using a custom programming script, which extracts latency timestamps for each electrode from the .xlsx files.
NOTE: This can be done manually. - In step 2.1.11, record the maximum number of threshold crossings and maximum time separation of threshold crossings for the one LFP waveform. Modify the script for inputting these LFP-defining parameters for each slice before running the script.
NOTE: Thresholding previously performed in analysis software clearly captures EAPs as a single event. However, LFPs are composed of a variable number of peaks depending on the shape of the waveform and the subsequent number of threshold crossings by the one event. - Modify the script to input the electrodes of interest before analysis.
- To determine synchronicity (defined in the script by modifiable time frames for synchronous activity to occur within), separate and analyze the extracted latencies to detect coincident events.
NOTE: The script allows the maximum time between coincident events to be set. These are set at 20 ms for EAPs and 200 ms for LFPs. - Run the script to extract latency timestamps.
NOTE: The .xlsx output file contains the interpretations of latency data, which are EAP and LFP counts, frequencies, and coincident event counts for individual electrodes and whole slices. These data are used to assess the frequency, EAP/LFP counts, number of active electrodes, number of coincident events, number of linked electrodes, and the average strength of these linkages.
- Perform data analysis using a custom programming script, which extracts latency timestamps for each electrode from the .xlsx files.
Model of network activity in the spinal cord dorsal horn
Application of 4-AP reliably induces synchronous rhythmic activity in the spinal cord DH. Such activity presents as increased EAPs and LFPs. The later signal is a low-frequency waveform, which has previously been described in MEA recordings30. Changes in EAP and/or LFP activity following drug application reflect altered neural activity. Examples of EAPs and LFPs are shown in Figure 3B and Figure 4. The focus here is on the following parameters or features of the EAP/LFP data: frequency, total counts, active electrode counts, synchronicity as characterized by the number of coincident events detected across multiple electrodes, number of linked adjacent electrodes, and the strength of linkages between adjacent electrodes. Representative results are shown in Table 3 and Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, and Figure 8. They show a significant increase in all the parameters measured (all p<0.001 by paired t-test or the Wilcoxon Signed-Rank non-parametric equivalent test) for both EAPs (Figure 5 and Figure 6) and LFPs (Figure 7 and Figure 8) following 4-AP stimulation and then relative stability for the remainder of the recordings. Data were tested for normality prior to statistical analysis. In summary, 4-AP induces EAP and LFP activity in the spinal cord DH, and various features of the data can be extracted from the MEA recordings. The activity is reproducible, and much of the activity, particularly for LFPs, is rhythmic and synchronous.
| Activity Feature | Baseline | 4-Aminopyridine | Significant Difference |
| Extracellular Action Potentials (EAPs) | |||
| Frequency | 0.07 ± 0.01 | 0.88 ± 0.09 | p<0.001 |
| Total Spike Count | 261.41 ± 70.62 | 3289. 57 ± 484.38 | p<0.001 |
| Active Electrode Count | 2.36 ± 0.34 | 8.95 ± 0.68 | p<0.001 |
| Number of Coincident Spikes | 9.26 ± 4.01 | 966.94 ± 189.21 | p<0.001 |
| Number of Linked Electrodes | 2.03 ± 0.42 | 24.06 ± 1.96 | p<0.001 |
| Strength of Linkages Between Electrodes | 1.97 ± 0.58 | 29.13 ± 4.60 | p<0.001 |
| Local Field Potentials (LFPs) | |||
| Frequency | 0.00 ± 0.00 | 0.28 ± 0.03 | p<0.001 |
| Total Spike Count | 4.79 ± 0.82 | 688.47 ± 121.16 | p<0.001 |
| Active Electrode Count | 0.41 ± 0.16 | 7.64 ± 0.73 | p<0.001 |
| Number of Coincident Spikes | 0.43 ± 0.23 | 108.06 ± 278.22 | p<0.001 |
| Number of Linked Electrodes | 0.24 ± 0.15 | 22.91 ± 2.46 | p<0.001 |
| Strength of Linkages Between Electrodes | 0.34 ± 0.19 | 29.20 ± 3.59 | p<0.001 |
Table 3: 4-Aminopyridine-induced activity. All presented as means ± SEM.
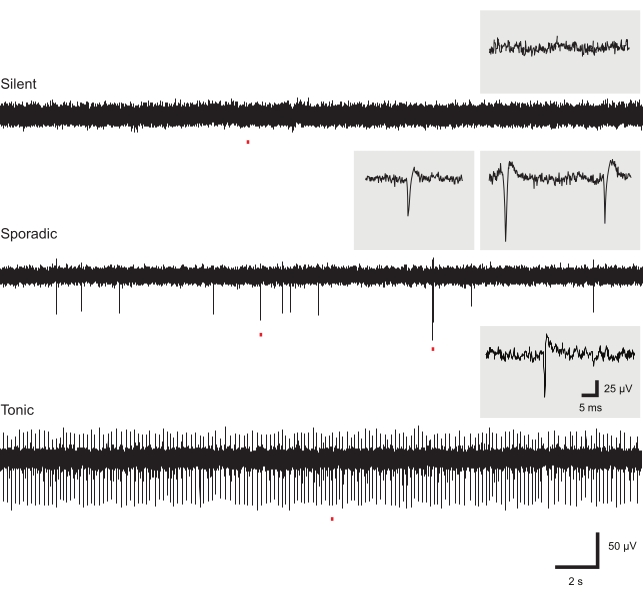
Figure 4: Exemplar baseline extracellular action potential activity. Panels show EAP activity (recordings are from different slices). Most electrodes in a given slice recording did not show baseline EAP activity (upper panel). Low-frequency sporadic EAPs were occasionally observed at baseline, potentially containing multiple spike waveforms (middle panel). High-frequency EAP activity was rarely observed in recordings at baseline (lower panel). Insets show individual EAPs from corresponding recordings on an expanded timescale. Abbreviation: EAP = extracellular action potential. Please click here to view a larger version of this figure.
Slice orientation
The DH circuitry activated by 4-AP is connected in all three dimensions. Thus, slice orientation is an important consideration for in vitro preparations. Sagittal or horizontal slicing may be preferential to observe intersegmental signaling, whereas transverse slices better preserve mediolateral and dorsoventral connectivity. Given these considerations, it can be seen that 4-AP stimulation induces similar rhythmic activity in the SDH, regardless of slice orientation (see Figure 9).
Long-term stability of 4-AP induced activity
The stability of 4-AP-induced activity is obviously crucial when studying the effects of applied drugs. Therefore, the stability of 4-AP-induced activity parameters was characterized, and this is presented in Figure 5, Figure 6, Figure 7, and Figure 8 and Table 4. All activity characteristics, plus the coincidence of activity for LFPs, were stable based on the similarity of 4-AP-induced activity at 12 min after 4-AP application and 15 min later (p>0.05). Other LFP synchronicity characteristics, the number of linked adjacent electrodes, and linkage strength between adjacent electrodes decreased over 15 min (p=0.016 and p=0.033, respectively), though the difference was modest. This gradual change could easily be distinguished from the more immediate actions of a test drug during pharmacological studies (see below). Data were tested for normal distribution before statistical comparisons and then assessed using paired t-tests or non-parametric Wilcoxon Signed-Rank tests as appropriate.
| Activity Feature | 4-Aminopyridine | 4-Aminopyridine (15 min) | Significant Difference |
| Extracellular Action Potentials (EAPs) | |||
| Frequency | 0.8 ± 0.13 | 0.85 ± 0.10 | p>0.05 (no dif.) |
| Total Spike Count | 2706.36 ± 510.96 | 2838.09 ± 447.73 | p>0.05 (no dif.) |
| Active Electrode Count | 9.32 ± 0.70 | 10.09 ± 0.56 | p>0.05 (no dif.) |
| Number of Coincident Spikes | 1037.63 ± 306.84 | 1013.09 ± 269.80 | p>0.05 (no dif.) |
| Number of Linked Electrodes | 22.00 ± 3.37 | 22.41 ± 2.56 | p>0.05 (no dif.) |
| Strength of Linkages Between Electrodes | 30.44 ± 6.27 | 31.88 ± 7.68 | p>0.05 (no dif.) |
| Local Field Potentials (LFPs) | |||
| Frequency | 0.25 ± 0.03 | 0.17 ± 0.03 | p>0.05 (no dif.) |
| Total Spike Count | 792.32 ± 155.83 | 546.32 ± 120.93 | p>0.05 (no dif.) |
| Active Electrode Count | 9.50 ± 1.11 | 7.86 ± 1.00 | p>0.05 (no dif.) |
| Number of Coincident Spikes | 1631.27 ± 734.77 | 1073.00 ± 490.85 | p>0.05 (no dif.) |
| Number of Linked Electrodes | 26.68 ± 4.58 | 20.95 ± 3.68 | p<0.05 |
| Strength of Linkages Between Electrodes | 33.35 ± 6.19 | 24.81 ± 5.41 | p<0.05 |
Table 4: 4-Aminopyridine activity stability. All presented as means ± SEM.
Pharmacological investigation of activity characteristics
To demonstrate that MEA-recorded 4-AP-induced activity is readily amenable to pharmacological manipulations, the dependence of these signals on action potential discharge was highlighted. Bath application of the voltage-gated sodium channel antagonist, tetrodotoxin (TTX, 1 μM), abolished both EAP and LFP activity, confirming spike dependency of these signals. Example traces are shown in Figure 3A. This result also provides an example of the utility of the preparation for future pharmacological investigations, where novel compounds and established analgesics can be assessed for their action in activated spinal DH circuits. Finally, to shed further light on the relevance of 4-AP activation of the DH networks, an alternative approach was trialed to achieve modest depolarization of the DH network. In this approach, an elevated potassium (4.5 mM) aCSF solution (Table 1) was bath-applied and shown to evoke a similar DH response to 4-AP stimulation. This manipulation evoked LFP activity that featured the same synchronous characteristics as 4-AP-induced responses (Figure 10), suggesting a similar mechanism and underlying circuitry.
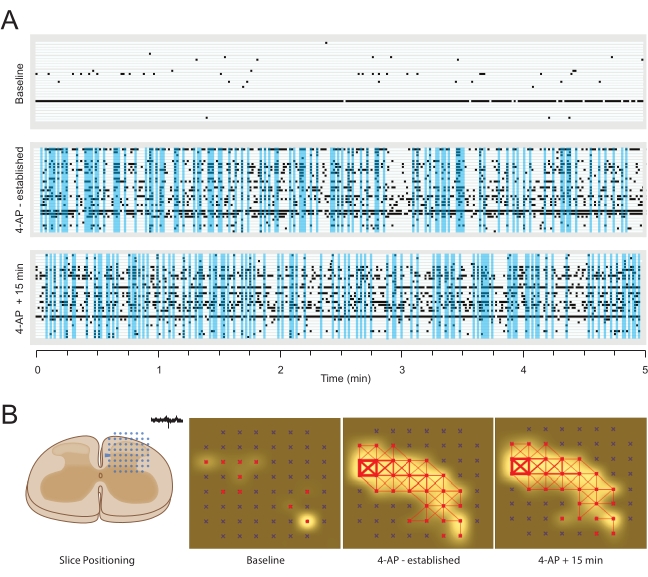
Figure 5: Example 4-aminopyridine-induced extracellular action potential activity. (A) Raster plots show EAP activity from active channels, detected at baseline (upper) and two timepoints (12 min – established, and 27 min) following bath addition of 4-AP (middle and lower). Vertical blue windows highlight periods of synchronous (close latency) activity in more than 5 recording electrodes. (B) Panels summarize EAP activity map analysis of MEA data. Left schematic shows the orientation of spinal cord slice relative to the electrode array. Middle left panel summarizes activity at baseline (active electrodes colored red) and EAP frequency indicated by white shading around active electrodes (shading intensity denotes increased activity). Middle right panel shows activity in the same slice after 12 min of 4-AP exposure. Note, the number of active electrodes (red) increased along with EAP frequency. In addition, synchrony between adjacent electrodes is indicated by red connecting lines, producing a network map of activity (line thickness denotes the degree of EAP similarity between electrodes). Right panel shows activity in the same slice following a further 15 min of 4-AP exposure. Note the number of active electrodes (red), degree of EAP activity (white), and network structure (red lines) have remained stable over this period. Abbreviations: 4-AP = 4-aminopyridine; EAP = extracellular action potential; MEA = microelectrode array. Please click here to view a larger version of this figure.
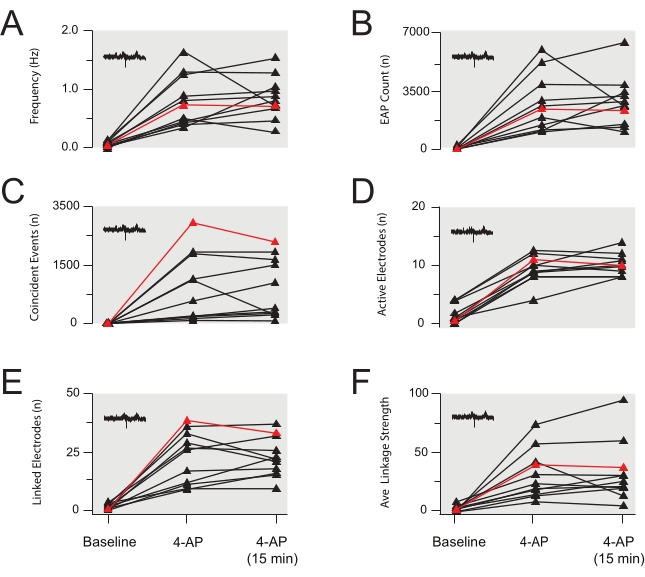
Figure 6: Group data summary of 4-aminopyridine-induced extracellular action potential activity. (A–F) Group data plots summarizing EAP properties from several experiments identical to the EAP data presented in Figure 4 (data also summarized in Table 3 and Table 4). EAP frequency (A), count (B), coincident events (C), active electrodes (D), linked electrodes (E), and average linkage strength (F) rose after bath application of 4-AP and were then stable for 15 min following the establishment of 4-AP-induced activity. Data are from 11 experiments (data in red is from the experiment in Figure 5). Abbreviations: 4-AP = 4-aminopyridine; EAP = extracellular action potential. Please click here to view a larger version of this figure.
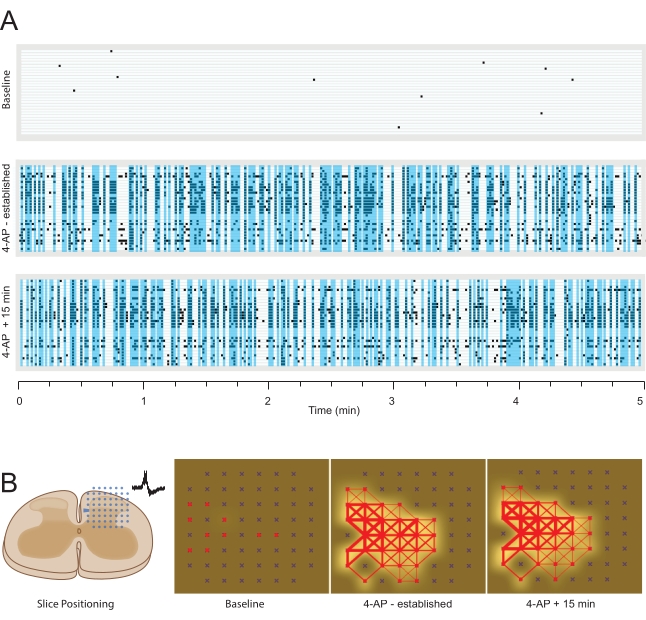
Figure 7: 4-Aminopyridine-induced local field potential activity. Data are presented as in Figure 5 except for LFP data. (A) Raster plots show LFP activity from multiple channels, detected at baseline (upper) and two timepoints (12 min – established, and 27 min) following bath addition of 4-AP (middle and lower). Vertical blue windows highlight periods of synchronous (close latency) activity in more than 5 recording electrodes. (B) Panels summarize LFP activity map analysis of MEA data. Left schematic shows the orientation of spinal cord slice relative to the electrode array. Middle left panel summarizes activity at baseline (active electrodes colored red), with minimal LFP frequency indicated by white shading around active electrodes (shading intensity denotes increased activity). Middle right panel shows activity in the same slice after 12 min of 4-AP exposure. The number of active electrodes (red) and LFP frequency are substantially increased. In addition, synchrony between adjacent electrodes (red connecting lines) shows a strong network map of LFP activity (line thickness denotes the degree of similarity between electrodes). Right panel shows LFP activity in the same slice following a further 15 min of 4-AP exposure. Note the number of active electrodes (red), degree of LFP activity (white), and network structure (red lines) are relatively stable over this period. Abbreviations: 4-AP = 4-aminopyridine; MEA = microelectrode array; LFP = local field potential. Please click here to view a larger version of this figure.
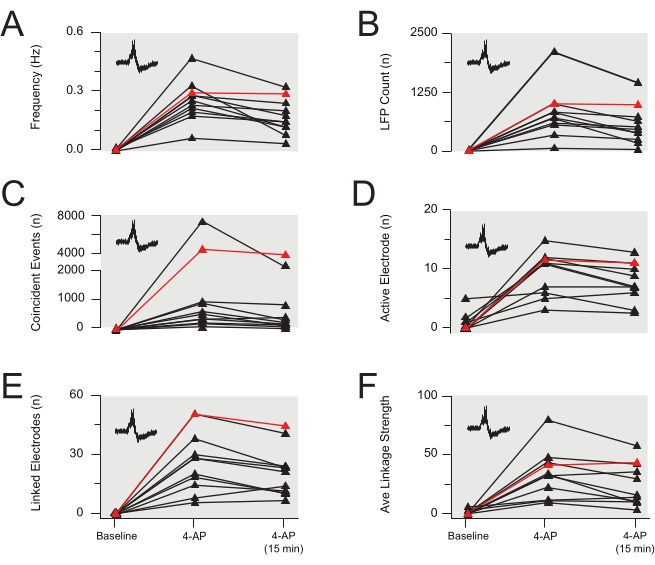
Figure 8: Group data summary of 4-aminopyridine-induced local field potential activity. (A–F) Group data plots summarizing LFP properties from several experiments identical to the EAP data presented in Figure 7 (data also summarized in Table 3 and Table 4). LFP frequency (A), count (B), coincident events (C), and active electrodes (D) were stable for 15 min after the 4-AP effect peaked (data in red is from the experiment in Figure 7). However, linked electrodes (E) and the average LFP linkage strength (F) decreased over time (both p<0.05). Data are from 11 experiments (data in red is from the experiment in Figure 7). Abbreviations: 4-AP = 4-aminopyridine; LFP = local field potential. Please click here to view a larger version of this figure.
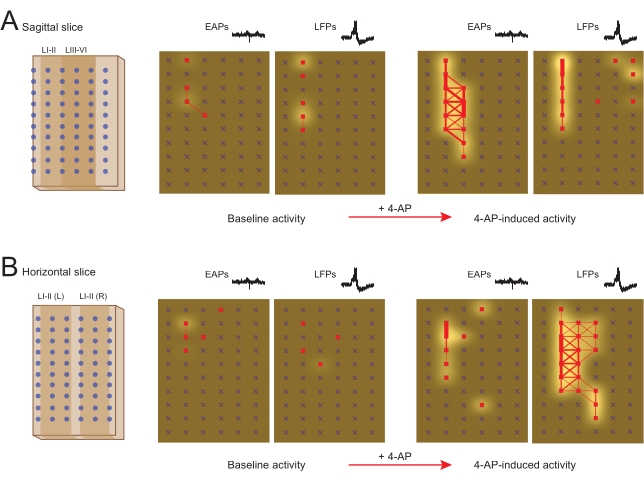
Figure 9: 4-Aminopyridine-induced extracellular action potential and local field potential activity in sagittal and horizontal slices. Panels summarize EAP and LFP activity in MEA network map analysis of 4-AP-induced signaling in sagittal (A) and horizontal (B) spinal cord slices. Schematics (far left) show the orientation of spinal cord slices relative to rectangular electrode arrays. Left network maps show baseline EAP and LFP activity in sagittal (A) and horizontal (B) spinal cord slices (active electrodes are red, frequency indicated by white shading intensity, and synchrony between adjacent electrodes by red connecting lines with thickness denoting the degree of synchrony). Right network maps show EAP and LFP activity in the same slice after 12 min of 4-AP exposure in sagittal (A) and horizontal (B) spinal cord slices. Note the substantial increase in the number of active electrodes, frequency of activity, and synchrony of these signals following 4-AP exposure, unmasking networks in both slice orientations. Abbreviations: MEA = microelectrode array; EAP = extracellular action potential; LFP = local field potential; 4-AP = 4-aminopyridine. Please click here to view a larger version of this figure.
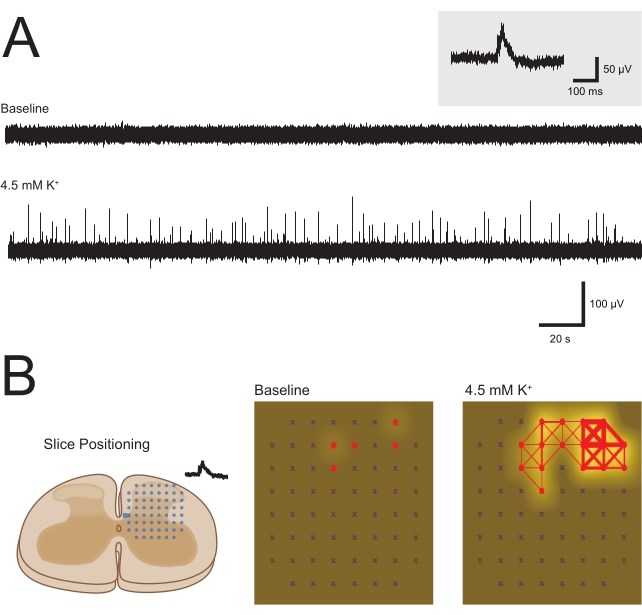
Figure 10: Elevated potassium (high K+)-induced local field potential activity. Panels summarize high K+ (4.5 mM) aCSF-induced LFP activity. (A) Example traces from one MEA channel at baseline and following bath addition of high K+ aCSF (5-min epochs). Elevation of K+ concentration produced clear LFP activity that was absent at baseline, similar to that seen with 4-AP application (Figure 3). Inset shows an LFP waveform on an expanded timescale. (B) Panels summarize LFP network activity induced by high K+ aCSF. Left schematic shows orientation of spinal cord slices relative to square electrode arrays. Network maps compare baseline and high K+-evoked LFP activity (active electrodes red, frequency indicated by white shading intensity, and synchrony between adjacent electrodes by red connecting lines with thickness denoting degree of synchrony). Note the substantial increase in the number of active electrodes, frequency of activity, and synchrony of these signals following high K+ aCSF exposure, unmasking the underlying network in a similar manner to 4-AP. Abbreviations: aCSF = artificial cerebrospinal fluid; MEA = microelectrode array; EAP = extracellular action potential; LFP = local field potential; 4-AP = 4-aminopyridine. Please click here to view a larger version of this figure.
