ORN axons arrive at the antennal lobe between 18 h and 36 h APF. They then navigate the antennal lobe, cross the midline, and innervate the glomeruli. Video 1 is a representative video showing the entire process for several individually identifiable axons, taken at the frequency of every 20 min for 24 h. Before registration using TurboReg, the axons exhibit some lateral drifting as the brain develops (first half of the video). After registration, the drifting is corrected (second half of the video).
To separate a few ORN axons from the same explant, one example is shown in Figure 5. Following the procedure in step 4.3, VA1d and VA1v axons from explant shown in Figure 5A were extracted to generate a new z stack image with only these two axons (Figure 5B). Similarly, VM2 and VM3 axons (Figure 5B') and DL2 axon (Figure 5B'') were extracted. Figure 5C shows a merge of images in Figure 5B-B'' with pseudocolors. The genetic identities of each ORN axons were revealed by immunostaining of a neuropil marker N-cadherin of the fixed explant (Figure 5D,E).
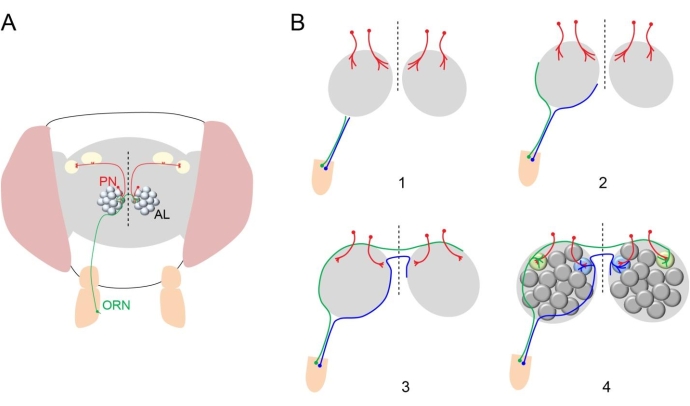
Figure 1: Structure of fly olfactory circuit. (A) An adult fly head is shown with one ORN from the right antennae (green) sending its axon to both antennal lobes (ALs) in the brain and forming synaptic connection in a specific glomerulus with dendrites of PNs (red) in the ipsilateral and contralateral ALs. Dashed vertical line indicates midline in this and subsequent diagrams and images. (B) Diagram showing the olfactory circuit development. (1) PN dendrites first innervate a region in the antennal lobe (red). ORN axons reach the antennal lobes in the brain. (2) ORN axons take either a dorsolateral (green) or ventromedial (blue) trajectory to circumnavigate the antennal lobe. (3) ORN axons cross the midline. (4) ORN axons innervate glomeruli in the antennal lobe. Please click here to view a larger version of this figure.

Figure 2: Preparation of the imaging chamber for the explant. (A) Lay a layer of silicone elastomer (~0.5 cm) at the bottom of a 60 mm Petri dish. (B–B') Align pins on a tape and cut to ~2 mm long using a pair of scissors. (C) Pin the micro pins on the silicone elastomer layer of the culture plate. Please click here to view a larger version of this figure.
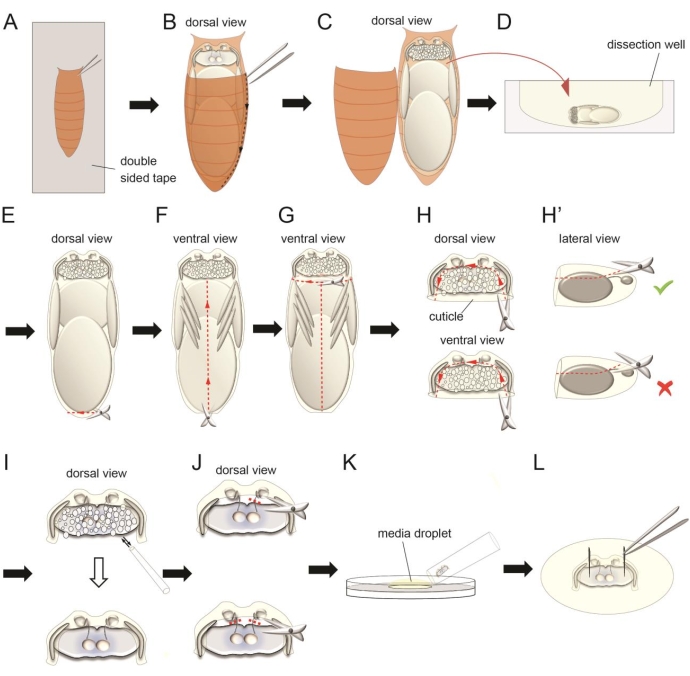
Figure 3: The dissection procedure for the antennae-brain explant. (A) Attach the ventral side of a paper tissue-dried pupa on a double-sided tape on a glass slide. (B–C) Use forceps to cut the external brown cuticle to expose the pupa inside. (D) Transfer the pupa to a dissection well with oxygenated full medium. (E–G) Carefully separate the pupal trunk from the head using microscissors. (H) Cut the pieces of semitransparent cuticle covering the dorsal and ventral sides of the brain. Keep some cuticle on the anterior and lateral sides of the brain to retain connections between the retina, antennae, and brain. (H') Avoid severing the antennal nerve during this step. (I) Clean the fat body covering the brain by gently pipetting. (J) Sever one or two antennal nerve(s) using microscissors in certain experiments. (K) Place a droplet of oxygenated full medium on the surface of the culture plate. Transfer the dissected explant using a wide tip pipette tip. (L) Use forceps to pin the two optic lobes of the explant on the silicone elastomer layer. Please click here to view a larger version of this figure.
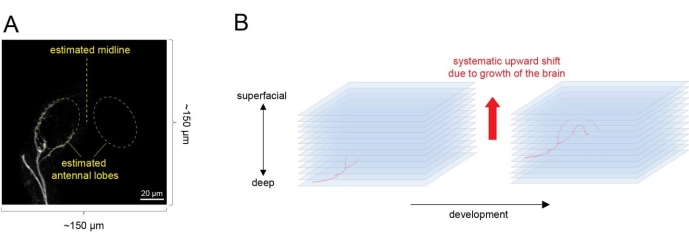
Figure 4: Time-lapse imaging of single ORN axon targeting from an explant. (A) Select an explant with a few ORN axons just reaching the antennal lobe. Estimate the shape of two antennal lobes by the curvature of the axons and center the antennal lobes in the imaging field. (B) Set the imaging region along the z axis. Consider that the antennal lobe will shift upward as the brain grows and develops. Please click here to view a larger version of this figure.
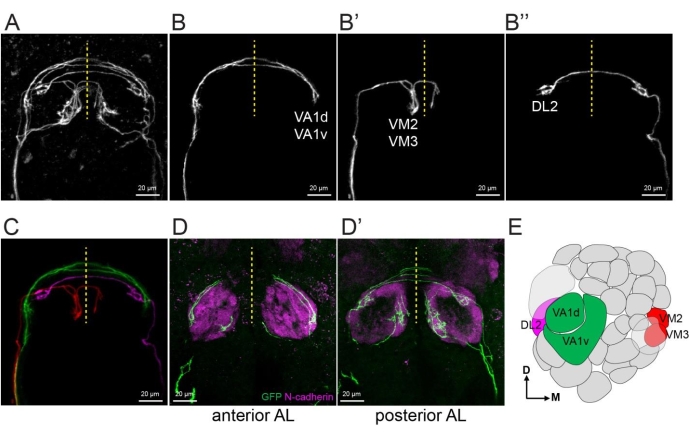
Figure 5: Extract single ORNs and reveal their glomerular identities. (A) A maximum projection image of an explant with 5-10 single ORN axons using two-photon microscopy with 20x objective and 3x zoom in. (B–B'') 1-2 single axons are extracted from (A) by manually creating masks in image sections from the raw image data. (C) Merge the images of (B-B'') with each axon pseudo-colored differently. (D–D') Maximum projection confocal images taken with 40x objective and 1.5x zoom. Explant shown in (A) was fixed followed by staining with anti-GFP and anti-N-cadherin (neuropil marker). Anterior and posterior halves of the antennal lobes are stacks separately in (D) and (D'). (E) Antennal lobe map shows extracted ORN axons in (B,C). Some images shown in this figure are modified from a prior study8. Please click here to view a larger version of this figure.
Video 1: Two-photon microscopy based time-lapse images show targeting of two ORN axons, before and after image registration. Please click here to download this Video.
