Les particules de type viral (VLP) ressemblent à la structure native des particules virales tout en manquant du génome viral, offrant ainsi un profil de sécurité élevé1,2. Les VLP représentent une classe individuelle de vaccins de plus en plus développés en raison de leur immunogénicité élevée3,4,5,6,7. C’est particulièrement le cas pour les VLP enveloppés de membrane, permettant l’affichage non seulement d’antigènes de surface viraux homologues, mais aussi d’antigènes hétérologues tels que les antigènes tumoraux8,9,10. La figure 1 donne un aperçu exemplaire de la structure d’un VLP enveloppé décoré d’antigènes. Au cours du processus de développement des vaccins à base de VLP, des tests sont indispensables pour permettre l’analyse de l’antigène cible respectif affiché sur la surface du VLP. De tels essais devraient être essentiels pour élucider la composition d’un vaccin particulaire: (i) Les VLP sont-ils décorés avec l’antigène de surface respectif? (ii) L’antigène de surface a-t-il conservé sa structure native comme l’a démontré la reconnaissance épitopique des anticorps neutralisants (bNAbs) et (iii) l’intégrité structurelle des VLP peut-elle être confirmée en raison de la détection de la protéine virale médiant la formation de VLP?
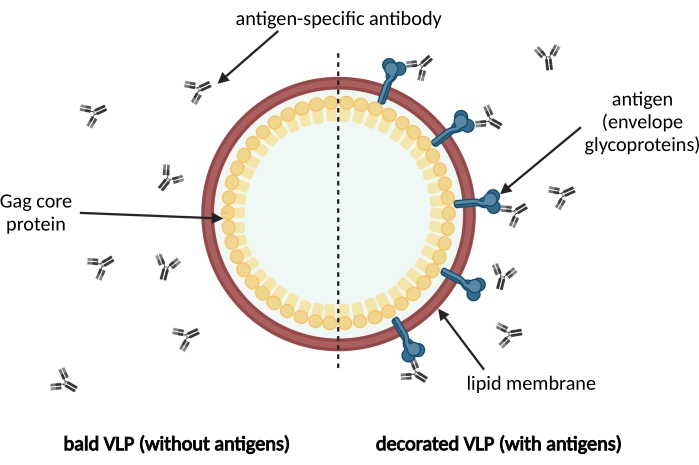
Figure 1 : Illustration schématique d’un VLP enveloppé d’une membrane. Les VLP sont formés par des protéines de noyau Gag précurseurs immatures et entourés d’une membrane lipidique dérivée de la cellule hôte. Les antigènes, par exemple les glycoprotéines d’enveloppe, sont incorporés dans la membrane lipidique et affichés à la surface du VLP (à droite). Les anticorps spécifiques de l’antigène reconnaissent l’antigène. Sur la gauche, un VLP chauve sans décoration antigénique est montré. Veuillez cliquer ici pour voir une version agrandie de cette figure.
En particulier, les VLP formés par la protéine précurseur de base p55 de l’antigène spécifique du groupe viral (Gag) du virus de l’immunodéficience humaine de type 1 (VIH-1) sont des échafaudages préférés pour l’affichage de l’antigène dans le développement de vaccins, car de nombreux anticorps et des kits ELISA sont disponibles, permettant la quantification de ces VLP11,12. Les glycoprotéines de l’enveloppe DU VIH-1 (Env), à savoir la protéine transmembranaire gp41 (gp41-TM) et l’unité de surface soluble gp120 (gp120-SU) formant des hétérodimères, sont incorporées dans l’enveloppe membranaire des particules et sont des antigènes cibles cruciaux pour le développement de vaccins contre l’infection par le VIH13,14,15 . L’affichage d’épitopes sensibles à la neutralisation dans ces antigènes cibles est une condition préalable pour provoquer une réponse anticorps largement neutralisante chez les vaccinés. Outre une réponse des lymphocytes T dirigée contre les protéines Gag, celle-ci est considérée comme un corrélat important de la protection contre l’infection par le VIH16. Par conséquent, et lors de la conception et de la production de VLP décorés d’antigènes candidats cibles, l’analyse ultérieure de la qualité des antigènes affichés représente une étape critique dans le processus de développement d’un vaccin.
L’immunoprécipitation (IP) est une technique largement utilisée pour la détection des interactions protéine-protéine et la purification de complexes protéiques à petite échelle17. Barret et al. a fait l’objet d’un premier rapport sur le développement de la propriété intellectuelle en 1960, mais cette méthode a été constamment améliorée. L’IP permet la capture et l’isolement d’un antigène cible (proie) d’une solution en utilisant un anticorps spécifique à l’antigène (appât) immobilisé par couplage à des billes18,19. Dans ce protocole, nous démontrons une variante de l’application IP classique en utilisant des VLP p55 formés par bâillon enveloppés de membrane comme proies et des bNAbs qui reconnaissent les épitopes sensibles à la neutralisation dans les protéines d’enveloppe affichées à la surface des VLP comme protéines d’appât. L’application réussie de ce test de capture VLP facilite la prédiction de la capacité des VLP à l’antigène positif testé à provoquer une réponse neutralisante des cellules B chez les personnes vaccinées. De telles propriétés immunogènes des candidats vaccins à base de VLP sont fréquemment démontrées dans de petits modèles animaux20,21,22.
Afin d’évaluer la qualité du nouveau candidat vaccin VLP, les tests de capture VLP ont été utilisés avec succès5,23,24. Cependant, le nombre de méthodes publiées est limité. Le test de capture VLP présenté ici commence par l’immobilisation de bNAbs spécifiques à Env sur des billes conjuguées à la protéine G, qui se lient à la région Fc des anticorps dérivés de mammifères. Les matrices typiques pour l’immobilisation de l’anticorps de choix sont l’agarose ou les billes magnétiques. Cependant, les billes magnétiques sont favorables aux applications à haut débit25. Dans l’étape suivante, les VLP affichant l’antigène cible sont capturés par des billes recouvertes de bNAb. Les complexes immunitaires formés constitués de VLP Env-positifs et de bNAbs immobilisés sont facilement enrichis à l’aide d’un aimant. Les complexes immunitaires isolés sont élués dans la dernière étape. Par la suite, les VLP peuvent être caractérisés biochimiquement. Ici, nous avons effectué une analyse par transfert Western en utilisant des anticorps spécifiques à la protéine de base virale p55 Gag pour démontrer que les antigènes Env cibles précipités abritaient non seulement les épitopes sensibles à la neutralisation, mais étaient également affichés sur les VLP formés par Gag. De plus, la détection des protéines Gag du noyau viral augmente la sensibilité du test de capture puisque les protéines Gag sont plus abondantes que les protéines Env dans un VLP. Dans le VIH-1, les protéines Env ne sont présentes qu’à un nombre à un ou deux chiffres26, alors que plus de 3 500 molécules Gag forment le noyau d’une particule27.
Comparé à d’autres techniques d’examen des interactions protéine-protéine28,29, le test de capture VLP offre une méthode alternative pour les laboratoires de recherche n’ayant pas accès à des instruments d’analyse coûteux. Par exemple, l’analyse microscopique électronique en transmission (TEM), la spectroscopie par résonance plasmonique de surface (SPR) et l’analyse de suivi des nanoparticules (NTA) peuvent être coûteuses. Le test de capture présenté ici permet également de soumettre ultérieurement des échantillons de VLP à antigène positif capturés pour une caractérisation plus poussée des protéines, par exemple en utilisant l’électrophorèse sur gel, l’immunoblotting, la microscopie électronique et la spectrométrie de masse (SEP), respectivement. Étant donné que la structure native de l’antigène cible est préservée pendant le test de capture VLP, la performance d’un PAGE natif et des techniques d’immunobuvardage ultérieures peut également être utilisée.
Le test de capture VLP représente une méthode facile à utiliser et sensible pour examiner la décoration des VLP avec des antigènes cibles exposant des épitopes sensibles à la neutralisation, et donc leur utilité en tant que futurs candidats vaccins.
