Viruslignende partikler (VLPer) ligner den opprinnelige viruspartikkelstrukturen mens de mangler viral genom, og gir dermed en høy sikkerhetsprofil1,2. VIP-er representerer en individuell klasse vaksiner som i økende grad utvikles på grunn av deres høye immunogenisitet3,4,5,6,7. Dette er spesielt tilfelle for membran-innhyllede VLPer, noe som gjør det mulig å vise ikke bare homologe virale overflateantigener, men også heterologe antigener som tumorantigener8,9,10. Figur 1 gir en eksemplarisk oversikt over strukturen til en innhyllet antigen-dekorert VLP. Under utviklingsprosessen av VLP-baserte vaksiner er analyser uunnværlige, noe som muliggjør analyse av det respektive målantigenet som vises på VLP-overflaten. Slike analyser bør være medvirkende til å belyse sammensetningen av en partikkelvaksine: (i) Er VLPene dekorert med det respektive overflateantigenet? (ii) Har overflateantigenet beholdt sin opprinnelige struktur som demonstrert ved epitopgjenkjenning av nøytraliserende antistoffer (bNAbs) og (iii) kan VLPs strukturelle integritet bekreftes på grunn av påvisning av det virale proteinmedierende VLP-formasjonen?
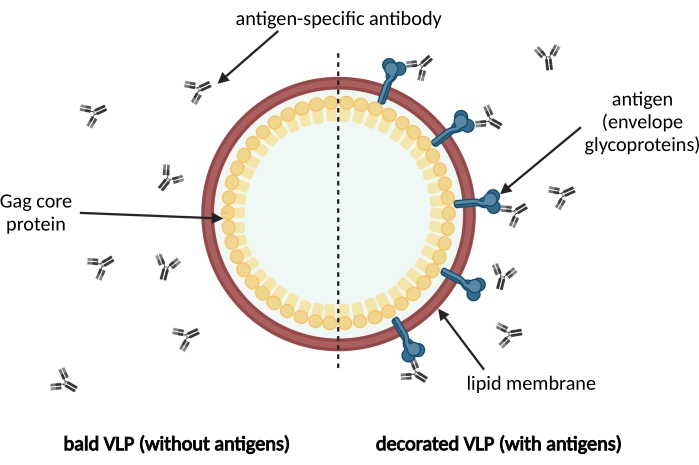
Figur 1: Skjematisk illustrasjon av en membran-innhylset VLP. VLPer dannes av umodne forløper Gag kjerneproteiner og er omgitt av en lipidmembran avledet fra vertscellen. Antigenene, for eksempel konvoluttglykoproteiner, er innlemmet i lipidmembranen og vises på overflaten av VLP (til høyre). Antigenspesifikke antistoffer gjenkjenner antigenet. Til venstre vises en skallet VLP uten antigendekorasjon. Klikk her for å se en større versjon av denne figuren.
Spesielt VLPer dannet av viral gruppespesifikk antigen (Gag) kjerneforløperprotein p55 av human immunsviktvirus type 1 (HIV-1) er foretrukne stillaser for antigenvisning i vaksineutvikling som mange antistoffer, og ELISA-sett er tilgjengelige, noe som muliggjør kvantifisering av disse VLPene11,12. HIV-1-konvoluttglykoproteinene (Env), nemlig transmembranproteinet gp41 (gp41-TM) og den oppløselige overflateenheten gp120 (gp120-SU) som danner heterodimere, er innlemmet i membrankonvolutten av partikler og er avgjørende målantigener for utvikling av vaksiner mot HIV-infeksjon13,14,15 . Visning av nøytraliseringsfølsomme epitoper i disse målantigenene er en forutsetning for å fremkalle en bredt nøytraliserende antistoffrespons hos vaksiner. Foruten en T-cellerespons rettet mot Gag-proteinene, regnes dette som en viktig korrelasjon av beskyttelse mot HIV-infeksjon16. Følgelig, og ved design og produksjon av VIP-er dekorert med målantigenkandidater, representerer den påfølgende analysen av kvaliteten på de viste antigenene et kritisk skritt i prosessen med vaksineutvikling.
Immunoprecipitation (IP) er en mye brukt teknikk for påvisning av proteinproteininteraksjoner og rensing av proteinkomplekser i liten skala17. Barret et al. først rapportert om utviklingen av IP i 1960, men denne metoden har blitt stadig ytterligere forbedret. IP muliggjør fangst og isolering av et målantigen (byttedyr) fra en løsning ved å bruke et antigenspesifikt antistoff (agn) immobilisert ved kobling til perler18,19. I denne protokollen demonstrerer vi en variasjon av den klassiske IP-applikasjonen ved hjelp av membrankonvolverte p55 Gag-dannede VLPer som byttedyr og bNAbs som gjenkjenner nøytraliseringsfølsomme epitoper i konvoluttproteinene som vises på overflaten av VLPene som agnproteiner. Den vellykkede anvendelsen av denne VLP-fangstanalysen letter forutsigelsen av om de testede antigenpositive VIP-ene vil kunne fremkalle en nøytraliserende B-cellerespons hos vaksinerte mennesker. Slike immunogene egenskaper hos VLP-baserte vaksinekandidater blir ofte demonstrert i små dyremodeller20,21,22.
For å vurdere kvaliteten på den nyutviklede VLP-vaksinekandidaten har VLP-fangstanalyser blitt brukt med hell5,23,24. Antallet publiserte metoder er imidlertid begrenset. VLP-fangstanalysen som presenteres her starter med immobilisering av Env-spesifikke bNAbs på protein G-konjugerte perler, som binder seg til Fc-regionen av pattedyr-avledede antistoffer. Typiske matriser for immobilisering av det valgte antistoffet er agarose eller magnetiske perler. Magnetiske perler er imidlertid gunstige for bruksområder med høy gjennomstrømning25. I neste trinn fanges VLPer som viser målantigenet av bNAb-belagte perler. De dannede immunkompleksene som består av Env-positive VIP-er og immobiliserte bNAbs, er lett beriket ved hjelp av en magnet. De isolerte immunkompleksene blir unngått i det siste trinnet. Deretter kan VLPene karakteriseres biokjemisk. Her utførte vi vestlig blotanalyse ved hjelp av p55 Gag virale kjerneproteinspesifikke antistoffer for å demonstrere at det utfelte målet Env antigener ikke bare inneholdt nøytraliseringsfølsomme epitoper, men ble også vist på Gag-dannede VLPer. Videre øker påvisning av viral kjerne Gag-proteiner følsomheten til fangstanalysen siden Gag-proteiner er mer rikelig enn Env i en VLP. I HIV-1 er Env-proteiner bare til stede på et enkelt- eller tosifret tall26, mens mer enn 3500 Gag-molekyler utgjør kjernen i en partikkel27.
Sammenlignet med andre teknikker for undersøkelse av proteinproteininteraksjoner28,29, gir VLP-fangstanalysen en alternativ metode for forskningslaboratorier som ikke har tilgang til dyre analytiske instrumenter. For eksempel kan transmisjonselektronmikroskopianalyse (TEM), overflateplasmonresonansspektroskopi (SPR) og nanopartikkelsporingsanalyse (NTA) være kostnadskrevende. Fangstanalysen som presenteres her, tillater også senere subjeksjon av fanget antigenpositive VLP-prøver for ytterligere proteinkarakterisering, for eksempel ved bruk av henholdsvis geleelektroforese, immunoblotting, elektronmikroskopi og massespektrometri (MS). Tatt i betraktning at den opprinnelige strukturen til målantigenet er bevart under VLP-fangstanalysen, kan også ytelsen til en innfødt SIDE og påfølgende immunoblottingsteknikker brukes.
VLP-fangstanalysen representerer en brukervennlig og sensitiv metode for å undersøke dekorasjonen av VIP-er med målantigener som eksponerer nøytraliseringsfølsomme epitoper, og dermed deres nytte som fremtidige vaksinekandidater.
