Optogenetikk, kontrollen av biologiske prosesser med lysresponsive proteiner, tilbyr en ny strategi for dynamisk styring av mikrobielle gjæringer for kjemisk og proteinproduksjon1,2. Byrden av konstruerte metabolske veier og toksisiteten til noen mellomprodukter og produkter svekker ofte celleveksten3. Slike påkjenninger kan føre til dårlig biomasseakkumulering og redusert produktivitet3. Denne utfordringen kan løses ved å dele gjæringer i en vekst- og produksjonsfase, som vier metabolske ressurser til henholdsvis biomasseakkumulering eller produktsyntese4. Vi viste nylig at overgangen fra vekst til produksjon i denne tofase gjæringen kan induseres med endringer i belysningsforholdene5,6,7. Den høye tunabiliteten, reversibiliteten og ortogonaliteten til lysinnganger8 gir unike fordeler for lyskontrollerte gjæringer som er vanskelige eller umulige å gjenskape med kjemiske indusere som brukes i dynamisk kontroll av konvensjonelle tofasede gjæringer4,9,10,11.
Det blålys responsive EL222-proteinet avledet fra Erythrobacter litoralis har blitt brukt til å utvikle flere optogenetiske kretser for metabolsk engineering i Saccharomyces cerevisiae5,7,12,13. EL222 inneholder et LOV-domene (light-oxygen-voltage sensor) som gjennomgår et konformasjonsskift ved aktivering av blått lys (465 nm), som gjør det mulig å binde seg til sin kojakk-DNA-sekvens (C120)13. Fusing EL222 til det virale VP16-aktiveringsdomenet (VP16-EL222) resulterer i en responsiv transkripsjonsfaktor med blått lys som kan aktivere genuttrykk i S. cerevisiae7 og andre organismer14 fra den syntetiske promotoren PC120. Flere kretser basert på EL222 er utviklet og brukt til kjemisk produksjon i S. cerevisiae, for eksempel det grunnleggende lysaktiverte OptoEXP-systemet7, der genet av interesse uttrykkes direkte fra PC120 (figur 1A). Imidlertid motiverte bekymringer for lysgjennomtrengning ved de høye celletetthetene som vanligvis oppstår i produksjonsfasen av gjæring oss til å utvikle inverterte kretser som er indusert i mørket, for eksempel OptoINVRT- og OptoQ-INVRT-kretsene (figur 1B) 5,7,13. Disse systemene utnytter galaktose (GAL) eller quinic acid (Q) reguloner fra henholdsvis S. cerevisiae og N. crassa, kontrollerer deres tilsvarende undertrykkere (GAL80 og QS) med VP16-EL222, for å undertrykke genuttrykk i lyset og sterkt indusere det i mørket. Ved å kombinere OptoEXP- og OptoINVRT-kretser resulterer det i toveis kontroll av genuttrykk, noe som muliggjør tofasede gjæringer der vekstfasen induseres med blått lys, og produksjonsfasen med mørke (figur 2A)5,7.
Å bruke lys i stedet for mørke for å indusere genuttrykk under produksjonsfasen vil i stor grad utvide evnene til optogenetiske kontroller, men vil også kreve å overvinne lysgjennomtrengningsbegrensningene til de høye celletetthetene som vanligvis oppstår i denne gjæringsfasen. For dette formål har vi utviklet kretser, kjent som OptoAMP og OptoQ-AMP, som forsterker transkripsjonsresponsen på blå lysstimulering. Disse kretsene bruker vilt- eller overfølsomme mutanter av VP16-EL222 for å kontrollere produksjonen av transkripsjonsaktivatorene Gal4p eller QF2 i henholdsvis GAL- eller Q-regulonene, og oppnår forbedret følsomhet og sterkere genuttrykk med lys12,13 (figur 1C). OptoAMP-kretser kan oppnå fullstendig og homogen lysinduksjon i 5 L bioreaktorer med optisk tetthet (målt ved 600 nm; OD600) verdier på minst 40 med bare ~ 0,35% belysning (5% lysdose på bare ~ 7% av bulkoverflaten). Dette viser en høyere grad av følsomhet sammenlignet med OptoEXP, som krever nær 100% belysning12. Evnen til effektivt å indusere genuttrykk med lys ved høye celletettheter åpner nye muligheter for dynamisk kontroll av gjæringer. Dette inkluderer driftsgjæringer i mer enn to tidsfaser, for eksempel trefasede gjæringer, der vekst, induksjon og produksjonsfaser etableres med unike lysplaner for å optimalisere kjemisk produksjon (figur 2B)12.
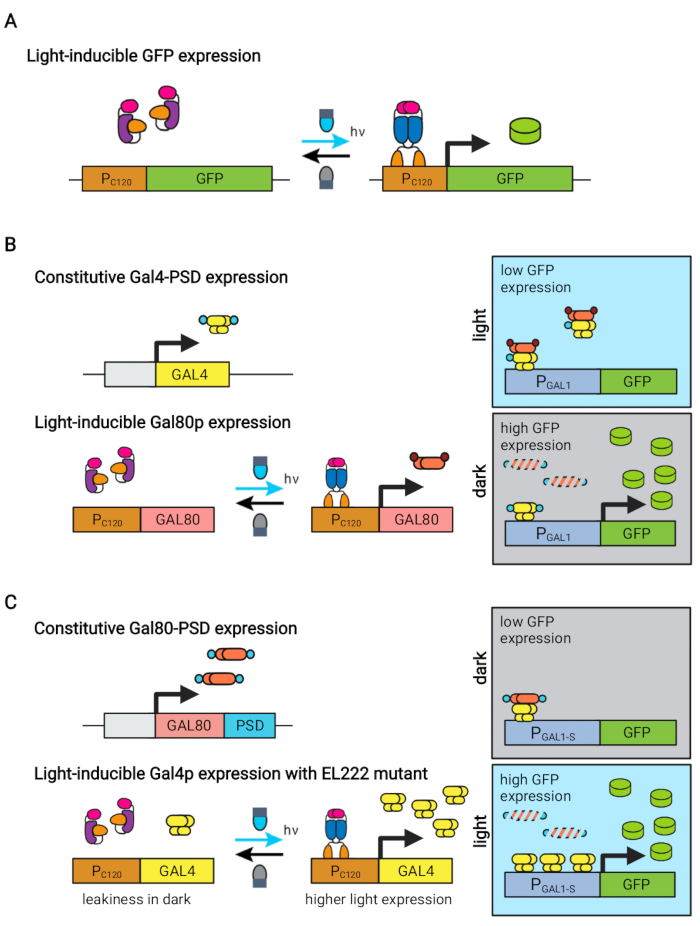
Figur 1: Optogenetiske kretser for dynamisk kontroll av S. cerevisiae. OptoEXP-, OptoINVRT- og OptoAMP-kretsene er basert på det lysfølsomme VP16-EL222-systemet. (A) I OptoEXP-kretsen forårsaker eksponering for blått lys en konformasjonsendring og dimerisering av VP16-EL222, som eksponerer et DNA-bindende domene og tillater transkripsjon fra PC120. Figuren er endret fra Zhao et al.7. (B) OptoINVRT-kretser utnytter GAL(vist) eller Q-reguloner for å indusere uttrykk i mørket. I GAL-baserte kretser uttrykkes VP16-EL222 og GAL4 konstituativt, mens PC120-stasjonsuttrykket til GAL80-repressoren (i Q-baserte kretser erstattes GAL4 og GAL80 av henholdsvis QF2 og QS, og en syntetisk QUAS-inneholdende promotor brukes i stedet for en GAL-promotor). I lys forhindrer Gal80p aktivering av genet av interesse fra PGAL1. I mørket uttrykkes ikke GAL80 og forringes raskt ved å fusjonere det til et konstituerende degrondomene (lite brunt domene), noe som muliggjør aktivering av PGAL1 av Gal4p. Figuren er endret fra Zhao et al.5. (C) OptoAMP-kretser bruker også VP16-EL222 til å kontrollere GAL- (vist) eller Q-reguloner. I disse kretsene er GAL80-repressoren (eller QS) konstituert uttrykt og smeltet sammen til en fotosensitiv degron (lite blått domene) som sikrer tett undertrykkelse i mørket. PC120 og en overfølsom VP16-EL222 mutant kontrolluttrykk av GAL4 (eller QF2) med lys, som sterkt aktiverer PGAL1 (eller en QUAS-inneholdende promotor) i lyset. GAL-avledede kretser kan bruke konstruerte former for PGAL1, for eksempel PGAL1-M eller PGAL1-S, som har økt aktivitet, samt wild-type promotorer kontrollert av GAL-regulonet (PGAL1, PGAL10, PGAL2, PGAL7). Figuren er endret fra Zhao et al.12. Klikk her for å se en større versjon av denne figuren.
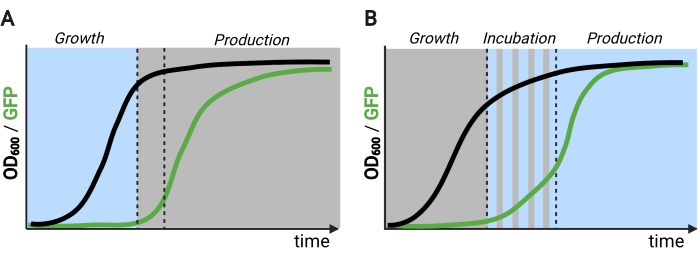
Figur 2: To- og trefasede gjæringer over tid. (A) Tofase gjæringer operert med inverterte kretser består av en lysdrevet vekstfase og en mørk produksjonsfase. I vekstfasen akkumuleres biomasse etter hvert som produksjonsveien forblir undertrykt. Når du når ønsket OD600, blir cellene flyttet til mørket for å justere metabolsk før de blir resuspendert i friske medier for produksjonsfasen. (B) I en trefaset prosess defineres vekst-, inkubasjons- og produksjonsfasene av unike lysplaner, som kan bestå av en mørk vekstperiode, pulsert inkubasjon og fullt opplyst produksjonsfase. Figur opprettet med Biorender. Klikk her for å se en større versjon av denne figuren.
Optogenetiske kretser er også utviklet for dynamisk kontroll av kjemisk og proteinproduksjon i E. coli. OptoLAC-kretser styrer den bakterielle LacI-repressoren ved hjelp av den lysresponsive pDawn-kretsen, som er basert på YF1/FixJ tokomponentsystem6 (figur 3). I likhet med OptoINVRT5 er OptoLAC-kretser designet for å undertrykke genuttrykk i lyset og indusere det i mørket. Uttrykksnivåer ved hjelp av OptoLAC-kretser kan matche eller overskride de som oppnås med standard isopropyl β-d-1-thiogalactopyranoside (IPTG) induksjon, og dermed opprettholde styrken av kjemisk induksjon samtidig som den tilbyr forbedret tunabilitet og reversibilitet6. Derfor muliggjør OptoLAC-kretser effektiv optogenetisk kontroll for metabolsk engineering i E. coli.
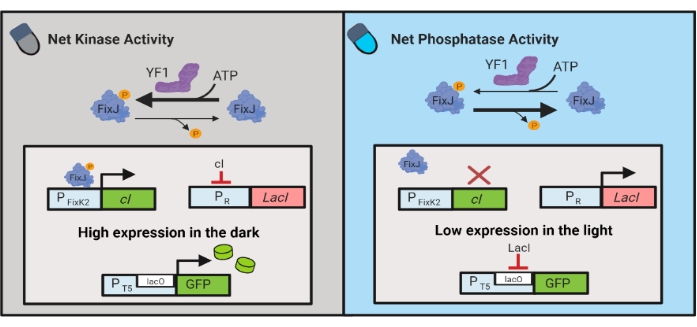
Figur 3: OptoLAC-kretser for dynamisk kontroll av E. coli. OptoLAC-kretsene tilpasser pDawn-systemet og lac operon for å oppnå aktivering i mørket og undertrykkelse i lyset. I mørket, YF1 fosforylater FixJ, som deretter aktiverer PFixK2 promotoren for å uttrykke cI-undertrykkeren . CI-undertrykkeren forhindrer uttrykk for laktI-undertrykkeren fra PR-promotoren , som tillater transkripsjon av genet av interesse fra en laktoholdig promotor. På den annen side reduserer blått lys YF1 netto kinase aktivitet, reversere FixJ fosforylering og dermed cI uttrykk, som derepresser uttrykk for lacI og forhindrer uttrykk fra lacO-inneholdende promotor. Figuren er endret fra Lalwani et al.6. Klikk her for å se en større versjon av denne figuren.
Vi beskriver her de grunnleggende protokollene for lyskontrollerte gjæringer av S. cerevisiae og E. coli for kjemisk eller proteinproduksjon. For både gjær og bakterier fokuserer vi først på gjæringer med en lysdrevet vekstfase og en mørkeindusert produksjonsfase aktivert av OptoINVRT- og OptoLAC-kretser. Deretter beskriver vi en protokoll for en trefaset (vekst, induksjon, produksjon) lyskontrollert gjæring aktivert av OptoAMP-kretser. Videre beskriver vi hvordan man skalerer opp optogenetisk kontrollerte gjæringer fra mikroplater til laboratorieskala bioreaktorer. Med denne protokollen tar vi sikte på å gi en komplett og lett reproduserbar guide for å utføre lyskontrollerte gjæringer for kjemisk eller proteinproduksjon.
