Metabolisme er den cellulære proces til produktion af energi. Cellulær metabolisme omfatter flere veje, herunder glykolyse, oxidativ fosforylering og glutaminolyse. Sunde celler bruger disse metaboliske veje til at generere energi til spredning og funktion, såsom produktion af cytokiner af immunceller. Mange sygdomme, herunder stofskifteforstyrrelser, kræft og neurodegeneration, er karakteriseret ved ændret cellulær metabolisme1. For eksempel har nogle kræftcelletyper forhøjede glykolysehastigheder, selv i nærvær af ilt, for at generere molekyler til syntese af nukleinsyrer, proteiner og lipider2,3. Dette fænomen, kendt som Warburg-effekten, er et kendetegn for mange kræfttyper, herunder brystkræft, lungekræft og glioblastomer4. På grund af ændringerne i cellulær metabolisme forbundet med kræftprogression kan cellulær metabolisme være en surrogatbiomarkør for lægemiddelrespons5,6. Desuden er forståelse af lægemiddeleffektivitet på celleniveau afgørende, da celleheterogenitet kan føre til forskellige lægemiddelresponser hos individer7,8.
Teknologier, der identificerer og kvantificerer ændringer i cellulær metabolisme, er afgørende for undersøgelser af kræft og lægemiddelrespons. Kemiske og proteinanalyser bruges til at evaluere metabolismen af celler eller væv, men mangler enkeltcelleopløsning og rumlig information. Metaboliske pladelæserbaserede assays kan måle pH og iltforbrug i prøven over tid og den efterfølgende metaboliske forstyrrelse af kemikalier. pH kan bruges til at beregne den ekstracellulære forsuringshastighed (ECAR), som giver et indblik i cellernes glykolytiske aktivitet9. Helkropsbilleddannelsesmetoder, herunder 2-[fluor-18] fluor-D-glucose positronemissionstomografi (FDG PET) og magnetisk resonansspektroskopi (MRS), er ikke-invasive billeddannelsesmetoder, der anvendes klinisk til at identificere tumorgentagelse og lægemiddeleffektivitet gennem metaboliske målinger10,11,12,13,14.
FDG-PET billeder vævsoptagelsen af FDG, en radioaktivt mærket glukoseanalog. Øget optagelse af FDG-PET af tumorer i forhold til omgivende væv skyldes Warburg-effekten12,13. MRS-billeder af almindelige kerner af molekyler, der anvendes til metabolisme, såsom 13C og 31P, og kan få dynamisk information om, hvordan stofskiftet ændres som reaktion på stimuli, såsom motion eller spisning14. Selvom FDG-PET og MRS kan bruges klinisk, mangler disse teknologier den rumlige opløsning til at løse intratumoral heterogenitet. Ligeledes foretages iltforbrugsmålinger på en bulkpopulation af celler. Autofluorescensbilleddannelse overvinder den rumlige opløsningshindring for disse teknologier og giver en ikke-invasiv metode til kvantificering af cellulær metabolisme.
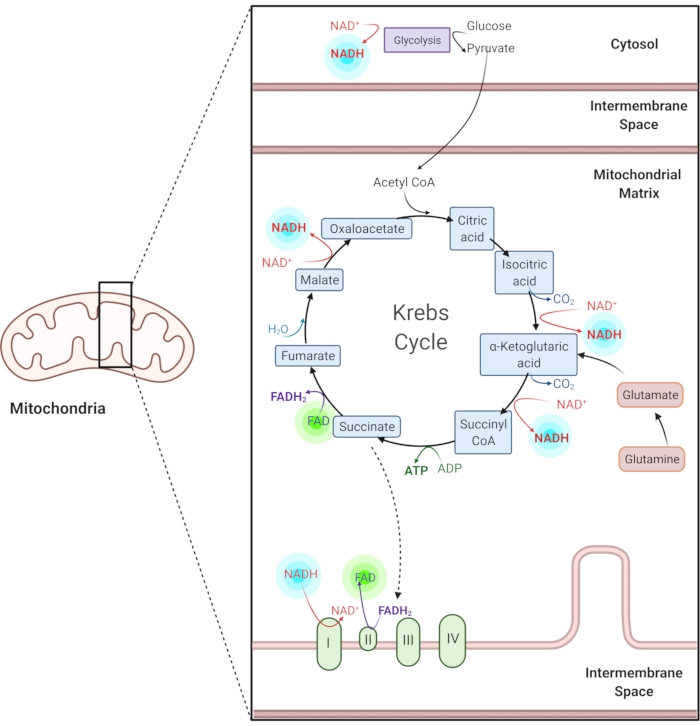
Figur 1: NADH og FAD i fælles metaboliske veje. NADH og FAD er coenzymer, der anvendes i glykolyse, Krebs-cyklussen og elektrontransportkæden. Autofluorescensbilleddannelse af disse molekyler giver information om cellulær metabolisme. Klik her for at se en større version af denne figur.
Reduceret nicotinamid adenin (phosphat) dinukleotid (NAD (P) H) og oxideret flavin adenin dinukleotid (FAD) er coenzymer af metaboliske reaktioner, herunder glycolyse, oxidativ phosphorylering og glutaminolyse (figur 1). Både NAD(P)H og FAD er autofluorescerende og giver endogen kontrast til fluorescensbilleddannelse1,15. NADPH har lignende fluorescerende egenskaber som NADH. På grund af dette bruges NAD (P) H ofte til at repræsentere det kombinerede signal fra NADH og NADPH2,16.
Fluorescens levetidsbilleddannelse (FLIM) kvantificerer fluorescensens levetid eller den tid, hvor en fluorofor er i ophidset tilstand. Fluorescensens levetid reagerer på fluoroforernes mikromiljø og giver information om cellulær metabolisme17. NAD(P)H og FAD kan eksistere i celler i enten proteinbundne eller frie konformationer, som hver især har en anden levetid. Fri NAD(P)H har en kortere levetid end proteinbundet NAD(P)H; Omvendt har fri FAD en længere levetid end bundet FAD18,19. Levetiden og levetidskomponentvægtene kan kvantificeres ud fra fluorescensens levetidshenfaldsdata gennem Eq. (1)20:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) repræsenterer den normaliserede fluorescensintensitet som funktion af tiden. De α 1 og α 2 i denne ligning repræsenterer de proportionale komponenter i korte og lange levetider (α 1+ α 2=1), henholdsvis τ1 og τ2 repræsenterer henholdsvis den korte og lange levetid, og C tegner sig for baggrundslys7,20. Den amplitudevægtede levetid, her repræsenteret som τm, beregnes ved hjælp af Eq. (2).
τm= α 1τ1+ α 2τ2 (2)
En gennemsnitlig levetid kan beregnes ved at beregne gennemsnittet af “t” over fluoroforens intensitetshenfald, som for et to-eksponentielt henfald er vist ved Eq. (3)17,21.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Et fluorescensintensitetsbillede kan beregnes ud fra livstidsbilledet ved at integrere fluorescensens levetidshenfald. Autofluorescensbilleddannelse er en ikke-destruktiv og etiketfri metode, der kan bruges til at karakterisere metabolismen af levende celler ved en subcellulær opløsning. Det optiske redoxforhold tilvejebringer en optisk analog metrik af cellens kemiske redoxtilstand og beregnes som forholdet mellem NAD (P) H og FAD intensiteter. Selvom formlen til beregning af det optiske redoxforhold ikke er standardiseret22,23,24,25, defineres den her som intensiteten af FAD over de kombinerede intensiteter af NAD (P) H og FAD. Denne definition anvendes, fordi den opsummerede intensitet i nævneren normaliserer metrikken mellem 0 og 1, og det forventede resultat af cyanidhæmningen er et fald i redoxforholdet. Fluorescensleveterne for fri NAD(P)H og FAD giver indsigt i ændringer i det metaboliske opløsningsmiddelmikromiljø, herunder pH, temperatur, nærhed til ilt og osmolaritet17.
Ændringer i fluorescenslevetid for de bundne fraktioner af NAD(P)H og FAD kan indikere udnyttelse af metabolisk vej og substratspecifik metabolisme26. Komponentvægte kan fortolkes for ændringer i den frie til den bundne brøkdel af coenzymerne18,19. Alt i alt tillader disse kvantitative autofluorescenslevetidsmålinger analyse af cellulær metabolisme, og autofluorescensbilleddannelse er blevet anvendt til at identificere neoplasmer fra normalt væv27,28, karakterisere stamceller29,30, evaluere immuncellefunktion31,32,33,34,35, måle neurologisk aktivitet36, 37,38 og forståelse af lægemiddeleffektivitet i kræfttyper som brystkræft og hoved- og halskræft21,39,40,41,42. Autofluorescensbilleddannelse i høj opløsning kan kombineres med billedsegmentering til enkeltcelleanalyse og kvantificering af intrapopulationsheterogenitet43,44,45,46,47.
NAD(P)H og FAD kan afbildes på enkeltfoton- eller multifotonfluorescensmikroskoper, der er konfigureret til intensitets- eller levetidsbilleddannelse. For enkeltfotonmikroskoper er NAD(P)H og FAD typisk spændte ved bølgelængder på henholdsvis 375-405 nm og 488 nm på grund af almindelige laserkilder ved disse bølgelængder48. I to-fotonfluorescens excitation vil NAD (P) H og FAD ophidse ved bølgelængder på henholdsvis ca. 700 til 750 nm og 700 til 900 nm15,49. Når fluoroforerne er spændte, udsender NAD (P) H og FAD fotoner ved bølgelængder mellem ~ 410 nm til ~ 490 nm og ~ 510 nm til ~ 640 nm, henholdsvis15. NAD(P)H- og FAD maxima-emissionsbølgelængderne er henholdsvis ca. 450 nm og 535 nm48.
På grund af deres forskellige excitations- og emissionsbølgelængder kan fluorescensen af de to metaboliske coenzymer isoleres spektralt. En forståelse af de spektrale egenskaber ved NAD (P) H og FAD er nødvendig for design og optimering af autofluorescensbilleddannelsesprotokoller. Cyanid er en elektrontransportkæde (ETC) kompleks IV-hæmmer. Virkningerne af cyanid på cellulær metabolisme og autofluorescensintensiteterne og levetiderne for NAD (P) H og FAD i celler er godt karakteriseret27,40. Derfor er et cyanidforstyrrelseseksperiment et effektivt middel til validering af NAD (P) H- og FAD-billeddannelsesprotokoller. Et vellykket cyanideeksperiment giver tillid til, at NAD (P) H og FAD-billeddannelsesprotokollen kan bruges til at vurdere metabolismen af ukendte grupper eller forstyrrelser.
