Metabolisme is het cellulaire proces van het produceren van energie. Cellulair metabolisme omvat meerdere routes, waaronder glycolyse, oxidatieve fosforylering en glutaminolyse. Gezonde cellen gebruiken deze metabole routes om energie te genereren voor proliferatie en functie, zoals de productie van cytokines door immuuncellen. Veel ziekten, waaronder metabole stoornissen, kanker en neurodegeneratie, worden gekenmerkt door een veranderd cellulair metabolisme1. Sommige soorten kankercellen hebben bijvoorbeeld verhoogde glycolysesnelheden, zelfs in de aanwezigheid van zuurstof, om moleculen te genereren voor de synthese van nucleïnezuren, eiwitten en lipiden2,3. Dit fenomeen, bekend als het Warburg-effect, is een kenmerk van vele soorten kanker, waaronder borstkanker, longkanker en glioblastomen4. Vanwege de veranderingen van het cellulaire metabolisme geassocieerd met kankerprogressie, kan cellulair metabolisme een surrogaatbiomarker zijn voor medicijnrespons5,6. Bovendien is het begrijpen van de werkzaamheid van geneesmiddelen op cellulair niveau cruciaal, omdat celheterogeniteit kan leiden tot verschillende geneesmiddelresponsen bij individuen7,8.
Technologieën die veranderingen in het cellulaire metabolisme identificeren en kwantificeren, zijn essentieel voor studies naar kanker en medicijnrespons. Chemische en eiwitanalyses worden gebruikt om het metabolisme van cellen of weefsels te evalueren, maar missen eencellige resolutie en ruimtelijke informatie. Metabole plaatlezer-gebaseerde assays kunnen de pH en het zuurstofverbruik in het monster in de loop van de tijd en de daaropvolgende metabole verstoring door chemicaliën meten. De pH kan worden gebruikt om de extracellulaire verzuringssnelheid (ECAR) te berekenen, wat inzicht geeft in de glycolytische activiteit van de cellen9. Beeldvormingsmethoden voor het hele lichaam, waaronder 2-[fluor-18] fluor-D-glucose positronemissietomografie (FDG PET) en magnetische resonantiespectroscopie (MRS), zijn niet-invasieve beeldvormingsmodaliteiten die klinisch worden gebruikt om tumorrecidief en werkzaamheid van geneesmiddelen te identificeren door middel van metabole metingen10,11,12,13,14.
FDG-PET brengt de weefselopname van FDG in beeld, een radioactief gelabeld glucose-analoog. Verhoogde opname van FDG-PET door tumoren ten opzichte van het omliggende weefsel is te wijten aan het Warburg-effect12,13. MRS toont gemeenschappelijke kernen van moleculen die worden gebruikt voor het metabolisme, zoals 13C en 31P, en kan dynamische informatie verkrijgen over hoe het metabolisme verandert als reactie op stimuli, zoals lichaamsbeweging of eten14. Hoewel FDG-PET en MRS klinisch kunnen worden gebruikt, missen deze technologieën de ruimtelijke resolutie om intratumorale heterogeniteit op te lossen. Evenzo worden metingen van het zuurstofverbruik uitgevoerd op een bulkpopulatie van cellen. Autofluorescentie beeldvorming overwint het ruimtelijke resolutie obstakel van deze technologieën en biedt een niet-invasieve methode voor het kwantificeren van cellulair metabolisme.
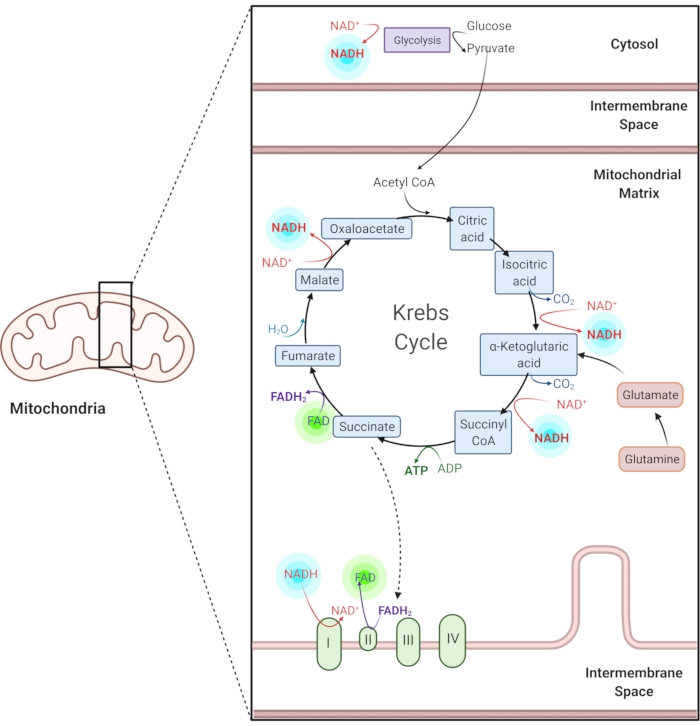
Figuur 1: NADH en FAD in gemeenschappelijke metabole routes. NADH en FAD zijn co-enzymen die worden gebruikt in glycolyse, de Krebs-cyclus en de elektronentransportketen. Autofluorescentie beeldvorming van deze moleculen geeft informatie over cellulair metabolisme. Klik hier om een grotere versie van deze figuur te bekijken.
Gereduceerd nicotinamide-adenine (fosfaat) dinucleotide (NAD (P)H) en geoxideerd flavine adenine dinucleotide (FAD) zijn co-enzymen van metabole reacties, waaronder glycolyse, oxidatieve fosforylering en glutaminolyse (figuur 1). Zowel NAD(P)H als FAD zijn autofluorescent en bieden endogene contrast voor fluorescentiebeeldvorming1,15. NADPH heeft vergelijkbare fluorescerende eigenschappen als NADH. Daarom wordt NAD(P)H vaak gebruikt om het gecombineerde signaal van NADH en NADPH2,16 weer te geven.
Fluorescence lifetime imaging (FLIM) kwantificeert de fluorescentielevensduur of de tijd waarvoor een fluorofoor zich in de aangeslagen toestand bevindt. Fluorescentielevensduur reageert op de micro-omgeving van de fluoroforen en geeft informatie over cellulair metabolisme17. NAD(P)H en FAD kunnen in cellen bestaan in eiwitgebonden of vrije conformaties, die elk een andere levensduur hebben. Vrije NAD(P)H heeft een kortere levensduur dan eiwitgebonden NAD(P)H; omgekeerd heeft vrije FAD een langere levensduur dan gebonden FAD18,19. De levensduur en het gewicht van de levensduurcomponenten kunnen worden gekwantificeerd op basis van gegevens over het levensduurverval van fluorescentie via Eq. (1)20:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) vertegenwoordigt de genormaliseerde fluorescentie-intensiteit als functie van de tijd. De α 1 en α 2 in deze vergelijking vertegenwoordigen de proportionele componenten van korte en lange levensduur (respectievelijk α 1+ α 2=1), respectievelijk τ1 en τ2 vertegenwoordigen respectievelijk de korte en lange levensduur, en C is goed voor achtergrondlicht7,20. De amplitudegewogen levensduur, hier weergegeven als τm, wordt berekend met eq. (2).
τm= α 1τ1+ α 2τ2 (2)
Een gemiddelde levensduur kan worden berekend door het gemiddelde van “t” over het intensiteitsverval van de fluorofoor, wat voor een twee-exponentieel verval wordt aangetoond door Eq. (3)17,21.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Een fluorescentie-intensiteitsbeeld kan worden berekend uit het levensduurbeeld door het fluorescentielevensverval te integreren. Autofluorescentie beeldvorming is een niet-destructieve en labelvrije methode die kan worden gebruikt om het metabolisme van levende cellen met een subcellulaire resolutie te karakteriseren. De optische redoxverhouding biedt een optische analoge metriek van de chemische redoxtoestand van de cel en wordt berekend als de verhouding van NAD(P)H- en FAD-intensiteiten. Hoewel de formule voor het berekenen van de optische redoxverhouding niet gestandaardiseerd is22,23,24,25, wordt deze hier gedefinieerd als de intensiteit van FAD over de gecombineerde intensiteiten van NAD(P)H en FAD. Deze definitie wordt gebruikt omdat de opgetelde intensiteit in de noemer de metriek tussen 0 en 1 normaliseert, en het verwachte resultaat van de cyanideremming een afname van de redoxverhouding is. De fluorescentielevensduur van vrije NAD(P)H en FAD geeft inzicht in veranderingen in de micro-omgeving van het metabole oplosmiddel, waaronder pH, temperatuur, nabijheid van zuurstof en osmolariteit17.
Veranderingen in de fluorescentielevensduur van de gebonden fracties van NAD(P)H en FAD kunnen wijzen op het gebruik van metabole routes en substraatspecifiek metabolisme26. Componentgewichten kunnen worden geïnterpreteerd voor veranderingen in de vrije tot de gebonden fractie van de co-enzymen18,19. Al met al maken deze kwantitatieve autofluorescentielevensduurmetingen de analyse van cellulair metabolisme mogelijk, en autofluorescentiebeeldvorming is gebruikt voor het identificeren van neoplasmata uit normale weefsels27,28, het karakteriseren van stamcellen29,30, het evalueren van de immuuncelfunctie31,32,33,34,35, het meten van neurologische activiteit36, 37,38, en inzicht in de werkzaamheid van geneesmiddelen bij kankersoorten zoals borstkanker en hoofd-halskanker21,39,40,41,42. Hoge-resolutie autofluorescentie beeldvorming kan worden gecombineerd met beeldsegmentatie voor eencellige analyse en kwantificering van intrapopulatie heterogeniteit43,44,45,46,47.
NAD(P)H en FAD kunnen worden afgebeeld op fluorescentiemicroscopen met één foton of multifotonen die zijn geconfigureerd voor intensiteits- of levenslange beeldvorming. Voor microscopen met één foton worden NAD(P)H en FAD meestal geëxciteerd op golflengten van respectievelijk 375-405 nm en 488 nm, als gevolg van gemeenschappelijke laserbronnen op deze golflengten48. Bij fluorescentie-excitatie van twee fotonen zullen NAD(P)H en FAD exciteren op golflengten van respectievelijk ongeveer 700 tot 750 nm en 700 tot 900 nm, 15,49. Zodra de fluoroforen zijn geëxciteerd, zenden NAD(P)H en FAD fotonen uit op golflengten tussen respectievelijk ~410 nm tot ~490 nm en ~510 nm tot ~640 nm15. De NAD(P)H en FAD maxima emissie golflengten zijn respectievelijk ongeveer 450 nm en 535 nm48.
Vanwege hun verschillende excitatie- en emissiegolflengten kan de fluorescentie van de twee metabole co-enzymen spectraal worden geïsoleerd. Een goed begrip van de spectrale kenmerken van NAD(P)H en FAD is noodzakelijk voor het ontwerp en de optimalisatie van autofluorescentie beeldvormingsprotocollen. Cyanide is een elektronentransportketen (ETC) complexe IV-remmer. De effecten van cyanide op het cellulair metabolisme en de autofluorescentie-intensiteiten en levensduur van NAD(P)H en FAD in cellen worden goed gekarakteriseerd27,40. Daarom is een cyanide perturbatie-experiment een effectief middel om NAD(P)H- en FAD-beeldvormingsprotocollen te valideren. Een succesvol cyanide-experiment geeft vertrouwen dat het NAD(P)H- en FAD-beeldvormingsprotocol kan worden gebruikt om het metabolisme van onbekende groepen of verstoringen te beoordelen.
