Le métabolisme est le processus cellulaire de production d’énergie. Le métabolisme cellulaire englobe de multiples voies, y compris la glycolyse, la phosphorylation oxydative et la glutaminolyse. Les cellules saines utilisent ces voies métaboliques pour générer de l’énergie pour la prolifération et la fonction, comme la production de cytokines par les cellules immunitaires. De nombreuses maladies, y compris les troubles métaboliques, le cancer et la neurodégénérescence, sont caractérisées par une altération du métabolisme cellulaire1. Par exemple, certains types de cellules cancéreuses ont des taux élevés de glycolyse, même en présence d’oxygène, pour générer des molécules pour la synthèse d’acides nucléiques, de protéines et de lipides2,3. Ce phénomène, connu sous le nom d’effet Warburg, est une caractéristique de nombreux types de cancer, y compris le cancer du sein, le cancer du poumon et les glioblastomes4. En raison des altérations du métabolisme cellulaire associées à la progression du cancer, le métabolisme cellulaire peut être un biomarqueur de substitution de la réponse aux médicaments5,6. De plus, la compréhension de l’efficacité des médicaments au niveau cellulaire est cruciale, car l’hétérogénéité cellulaire peut entraîner des réponses différentes aux médicaments chez les individus7,8.
Les technologies qui identifient et quantifient les changements dans le métabolisme cellulaire sont essentielles pour les études sur le cancer et la réponse aux médicaments. Les analyses chimiques et protéiques sont utilisées pour évaluer le métabolisme des cellules ou des tissus, mais manquent de résolution unicellulaire et d’informations spatiales. Les tests basés sur des lecteurs de plaques métaboliques peuvent mesurer le pH et la consommation d’oxygène dans l’échantillon au fil du temps et la perturbation métabolique subséquente par les produits chimiques. Le pH peut être utilisé pour calculer le taux d’acidification extracellulaire (ECAR), ce qui donne un aperçu de l’activité glycolytique des cellules9. Les méthodes d’imagerie du corps entier, y compris la tomographie par émission de positons 2-[fluor-18] fluoro-D-glucose (FDG PET) et la spectroscopie par résonance magnétique (MRS), sont des modalités d’imagerie non invasives utilisées cliniquement pour identifier la récidive tumorale et l’efficacité des médicaments par des mesures métaboliques10,11,12,13,14.
FDG-PET image l’absorption tissulaire de FDG, un analogue du glucose radiomarqué. L’absorption accrue de FDG-PET par les tumeurs par rapport aux tissus environnants est due à l’effet Warburg12,13. MRS image les noyaux communs des molécules utilisées pour le métabolisme, telles que le 13C et le 31P, et peut obtenir des informations dynamiques sur la façon dont le métabolisme change en réponse à des stimuli, tels que l’exercice ou l’alimentation14. Bien que fdG-PET et MRS puissent être utilisés cliniquement, ces technologies n’ont pas la résolution spatiale nécessaire pour résoudre l’hétérogénéité intratumorale. De même, les mesures de consommation d’oxygène sont effectuées sur une population en vrac de cellules. L’imagerie par autofluorescence surmonte l’obstacle de résolution spatiale de ces technologies et fournit une méthode non invasive de quantification du métabolisme cellulaire.
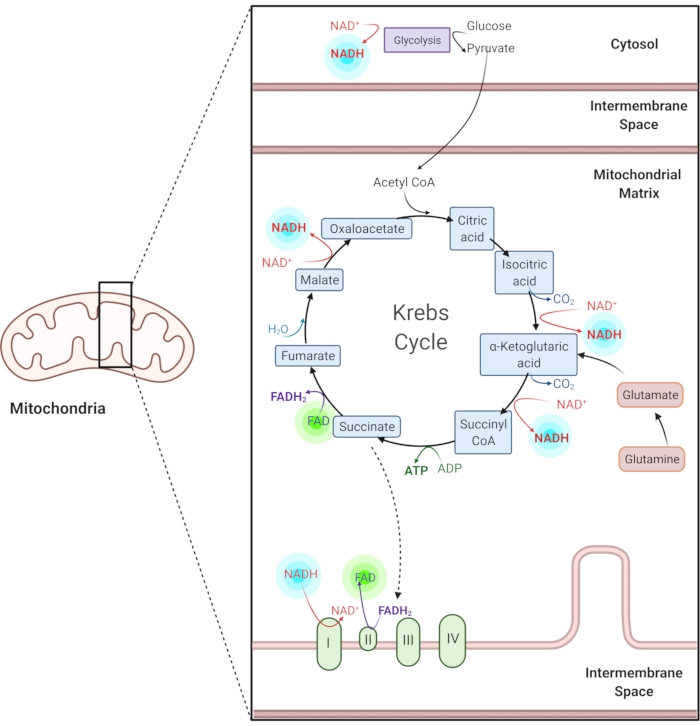
Figure 1 : NADH et FAD dans les voies métaboliques courantes. Le NADH et le FAD sont des coenzymes utilisées dans la glycolyse, le cycle de Krebs et la chaîne de transport d’électrons. L’imagerie par autofluorescence de ces molécules fournit des informations sur le métabolisme cellulaire. Veuillez cliquer ici pour voir une version agrandie de cette figure.
La nicotinamide adénine (phosphate) dinucléotide (NAD(P)H) réduite et la flavine adénine dinucléotide (FAD) oxydée sont des coenzymes de réactions métaboliques, y compris la glycolyse, la phosphorylation oxydative et la glutaminolyse (Figure 1). Le NAD(P)H et le FAD sont tous deux autofluorescents et fournissent un contraste endogène pour l’imagerie par fluorescence1,15. Le NADPH a des propriétés fluorescentes similaires à celles du NADH. Pour cette raison, nad(P)H est souvent utilisé pour représenter le signal combiné du NADH et du NADPH2,16.
L’imagerie de la durée de vie de la fluorescence (FLIM) quantifie la durée de vie de la fluorescence ou le temps pendant lequel un fluorophore est à l’état excité. Les durées de vie de la fluorescence sont sensibles au microenvironnement des fluorophores et fournissent des informations sur le métabolisme cellulaire17. Le NAD(P)H et le FAD peuvent exister dans les cellules dans des conformations liées aux protéines ou libres, chacune ayant une durée de vie différente. Le NAD(P)H libre a une durée de vie plus courte que le NAD(P)H lié aux protéines; à l’inverse, le FAD libre a une durée de vie plus longue que le FAD lié18,19. Les durées de vie et les poids des composants à vie peuvent être quantifiés à partir des données de désintégration de la durée de vie de la fluorescence via Eq. (1)20:
I(t) = α 1e-t/τ1 + α 2e-t/τ2 + C (1)
Eq (1) représente l’intensité de fluorescence normalisée en fonction du temps. Les α 1 et α 2 de cette équation représentent les composantes proportionnelles des durées de vie courtes et longues (α 1+ α 2=1), respectivement, τ1 et τ2 représentent les durées de vie courtes et longues, respectivement, et C représente la lumière de fond7,20. La durée de vie pondérée en fonction de l’amplitude, représentée ici par τm, est calculée à l’aide de Eq. (2).
τm= α 1τ1+ α 2τ2 (2)
Une durée de vie moyenne peut être calculée en faisant la moyenne de « t » sur la désintégration d’intensité du fluorophore, qui pour une désintégration à deux exponentielles est montrée par Eq. (3)17,21.
τ*m= (α 1τ12+ α 2τ22)/ (α 1τ1+ α 2τ2) (3)
Une image d’intensité de fluorescence peut être calculée à partir de l’image de durée de vie en intégrant la désintégration de la durée de vie de fluorescence. L’imagerie par autofluorescence est une méthode non destructive et sans étiquette qui peut être utilisée pour caractériser le métabolisme des cellules vivantes à une résolution subcellulaire. Le rapport redox optique fournit une métrique analogique optique de l’état redox chimique de la cellule et est calculé comme le rapport des intensités NAD(P)H et FAD. Bien que la formule de calcul du rapport redox optique ne soit pas normalisée22,23,24,25, elle est définie ici comme l’intensité de la DCP par rapport aux intensités combinées de NAD(P)H et de FAD. Cette définition est utilisée parce que l’intensité totale dans le dénominateur normalise la métrique entre 0 et 1, et le résultat attendu de l’inhibition du cyanure est une diminution du rapport redox. Les durées de vie en fluorescence du NAD(P)H libre et du FAD donnent un aperçu des changements dans le microenvironnement du solvant métabolique, y compris le pH, la température, la proximité de l’oxygène et l’osmolarité17.
Les changements dans la durée de vie de fluorescence des fractions liées de NAD(P)H et de FAD peuvent indiquer l’utilisation de la voie métabolique et le métabolisme spécifique du substrat26. Les poids des composants peuvent être interprétés pour les changements dans la fraction libre à la fraction liée des coenzymes18,19. Dans l’ensemble, ces mesures quantitatives de la durée de vie de l’autofluorescence permettent l’analyse du métabolisme cellulaire, et l’imagerie par autofluorescence a été utilisée pour identifier les néoplasmes des tissus normaux27,28, caractériser les cellules souches29,30, évaluer la fonction des cellules immunitaires31,32,33,34,35, évaluer l’activité neurologique36, 37,38, et comprendre l’efficacité des médicaments dans les types de cancer tels que le cancer du sein et le cancer de la tête et du cou21,39,40,41,42. L’imagerie par autofluorescence à haute résolution peut être combinée à la segmentation d’images pour l’analyse unicellulaire et la quantification de l’hétérogénéité intrapopulation43,44,45,46,47.
Le NAD(P)H et le FAD peuvent être imagés sur des microscopes à fluorescence monophotonique ou multiphotonique configurés pour l’imagerie d’intensité ou de durée de vie. Pour les microscopes monophotoniques, le NAD(P)H et le FAD sont généralement excités à des longueurs d’onde de 375-405 nm et 488 nm, respectivement, en raison de sources laser courantes à ces longueurs d’onde48. Dans l’excitation par fluorescence à deux photons, le NAD(P)H et le FAD s’exciteront à des longueurs d’onde d’environ 700 à 750 nm et de 700 à 900 nm, respectivement15,49. Une fois les fluorophores excités, le NAD(P)H et le FAD émettent des photons à des longueurs d’onde comprises respectivement entre ~410 nm et ~510 nm à ~640 nm15. Les longueurs d’onde d’émission maximales NAD(P)H et FAD sont respectivement d’environ 450 nm et 535 nm48.
En raison de leurs différentes longueurs d’onde d’excitation et d’émission, la fluorescence des deux coenzymes métaboliques peut être isolée spectralement. Une compréhension des caractéristiques spectrales du NAD(P)H et du FAD est nécessaire pour la conception et l’optimisation des protocoles d’imagerie par autofluorescence. Le cyanure est un inhibiteur IV du complexe IV de la chaîne de transport d’électrons (ETC). Les effets du cyanure sur le métabolisme cellulaire et les intensités d’autofluorescence et la durée de vie du NAD(P)H et du FAD dans les cellules sont bien caractérisés27,40. Par conséquent, une expérience de perturbation du cyanure est un moyen efficace de valider les protocoles d’imagerie NAD(P)H et FAD. Une expérience réussie sur le cyanure permet de s’assurer que le protocole d’imagerie NAD(P)H et FAD peut être utilisé pour évaluer le métabolisme de groupes inconnus ou de perturbations.
