قد يساعد نموذج الفأر للتنكس العصبي الذي تم تحليله هنا في دراسة العديد من الجوانب الحرجة المشاركة في الفيزيولوجيا المرضية لمرض باركنسون ، بما في ذلك الآليات المشاركة في علم أمراض αSyn والتنشيط الدبقي الصغير ، ومشاركة الجهاز المناعي المحيطي في تنظيم الالتهاب العصبي ، وآليات التنكس العصبي. من بين الآليات المشاركة في علم أمراض αSyn هي تلك الآليات دون الخلوية المرتبطة بخلل الميتوكوندريا أو الليزوسومات أو البروتيسومال في وجود حمل مفرط من αSyn في الخلايا العصبية الدوبامين في SN2. من المهم مراعاة أنه بالإضافة إلى تعبير hαSyn الناجم عن النقل بوساطة AAV ، يساهم الماوس الداخلي αSyn أيضا في حمل التعبير الكلي αSyn. الفئران المعدلة وراثيا التي تبالغ في التعبير عن الفئران αSyn تطور أمراض سينوكلين مماثلة ، وعلم الأمراض العصبية ، وضعف حركي لتلك النماذج الفئران على أساس الإفراط في التعبير عن hαSyn32. فيما يتعلق بالتنشيط الدبقي الصغير ، يمكن استخدام نموذج الفأر الحالي لدراسة كيف يمكن للاعبين الجزيئيين والخلويين المختلفين مثل السيتوكينات والناقلات العصبية والخلايا النجمية والخلايا العصبية والحاجز الدموي الدماغي والخلايا التائية تنظيم اكتساب الأنماط الظاهرية الوظيفية المؤيدة للالتهابات أو المضادة للالتهابات 8,10,11 . يشكل هذا النموذج أيضا أداة مهمة لدراسة دور الجهاز المناعي المحيطي ، بما في ذلك ليس فقط الخلايا التائية ولكن أيضا البلاعم والخلايا الوحيدة والعدلات ، على عمليات الالتهاب العصبي والتنكس العصبي للخلايا العصبية النيجيرية11،33،34. وأخيرا ، يمثل نموذج الفأر هذا أيضا نظاما قيما لدراسة الآليات الخلوية والجزيئية للتنكس العصبي في الجسم الحي ، بما في ذلك تلك التي تسببها العمليات الخلوية الداخلية ، مثل الإجهاد التأكسدي ، وعجز الطاقة ، والعضيات التالفة 2 ، أو تلك التي يمارسها اللاعبون الخارجيون ، مثل العوامل السمية العصبية التي تنتجها الخلايا الدبقية الدقيقة ، والخلايا النجمية ، والخلايا التائية السامة للخلايا8 ، 28,29,35.
أحد قيود نموذج الفأر هذا هو دراسة كيف أن التجميع المرضي ل αSyn في مواقع خارج الدماغ قد يشكل المراحل الأولية في تطور مرض باركنسون36. في هذا الصدد ، هناك أدلة متزايدة تشير إلى أنه قبل التنكس العصبي للخلايا العصبية النيجيرية والضعف الحركي ، يبدأ علم أمراض αSyn في الغشاء المخاطي للأمعاء والظهارة الشمية36 ، وربما استجابة الخلايا التائية الخاصة ب αSyn وكذلك12. بعد ذلك ، ستنتقل مجاميع αSyn عبر العصب المبهم إلى جذع الدماغ ، مما يؤدي إلى التهاب الأعصاب والتنكس العصبي للخلايا العصبية الدوبامينية12. على الرغم من أن نموذج AAV-hαSyn يلخص معظم جوانب مرض باركنسون ، إلا أنه لا توجد مشاركة واضحة للتجميع المرضي ل αSyn في مواقع خارج الدماغ في هذا النموذج. قد يكون النموذج البديل الذي يتضمن علم أمراض hαSyn مناسبا لدراسة هذه الجوانب من مرض باركنسون هو الفئران المعدلة وراثيا التي تبالغ في التعبير عن hαSyn تحت سيطرة مروج Thy1 ، نموذج Thy1-SNCA 37 ، حيث يعتمد تطور المرض على ميكروبات الأمعاء وينطوي على ضعف واضح في الجهاز الهضمي38.
على الرغم من أنه مفيد لدراسة العمليات المتنوعة المرتبطة بالفيزيولوجيا المرضية لمرض باركنسون ، إلا أن نموذج الفأر الحالي ينطوي على خطوات حاسمة يجب فحصها بدقة ، بما في ذلك التسليم الصحيح للناقلات الفيروسية في الإحداثيات المكانية المقابلة ، والتعبير الانتقائي عن hαSyn في الخلايا العصبية (التي تعتمد على النمط المصلي AAV وبناء المتجه) ، وجرعة AAV المناسبة وتوقيتها قبل تحليل النمط الظاهري لمرض باركنسون. من الضروري تحليل التوصيل السليم للناقلات الفيروسية في SN ، لأن استخدام الإحداثيات المكانية الصحيحة ل SN قد لا يكون كافيا عندما لا تكون الإبرة مستقيمة تماما ، والتي تكون في بعض الأحيان غير محسوسة للعين البشرية. علاوة على ذلك ، يعتمد انتشار ناقلات AAV على النمط المصلي AAV39. ولهذه الأسباب، من الضروري إجراء ضوابط دورية للجودة للتحقق من التسليم والانتشار الصحيحين لناقلات AAV-GFP المحقونة بعد ملاحظة GFP في شرائح الدماغ التي تحتوي على منطقة SN.
فيما يتعلق بالتعبير الانتقائي ل hαSyn في الخلايا العصبية ، من حيث المبدأ ، يمكن هندسة التعبير عن hαSyn ليتم التحكم فيه من قبل مروج انتقائي للخلايا العصبية أو ، بشكل أكثر دقة ، انتقائي للخلايا العصبية الدوبامينية ، مثل استخدام مروج TH في ناقلات AAV للحث على التعبير الانتقائي للجينات في الخلايا العصبية الدوبامينية40 . ومع ذلك ، فإن هذه الاستراتيجية لا تعمل عندما يكون ما يتم البحث عنه هو التعبير المفرط عن جين الاهتمام. لهذا السبب ، في النموذج الحالي ، من الضروري استخدام مروج قوي (مروج يحفز تعبيرا عاليا عن الجين النهائي) والأنماط المصلية AAV مع الاستوائية العصبية. في هذه الدراسة ، تم استخدام مروج CBA كمروج قوي للحث على الإفراط في التعبير عن hαSyn ، وتم استخدام النمط المصلي AAV5 للناقل الفيروسي. تم استخدام هذا النمط المصلي من قبل لنقل الخلايا العصبية للفئران والفئران41,42. هنا ، أظهرت النتائج أنه بعد 12 أسبوعا من تسليم AAV5-GFP في SN من الفئران ، كان التألق الأخضر موجودا بشكل انتقائي على الجانب الجانبي لكل من SN والمخطط (الشكل 1) ، مما يشير إلى النقل الفعال للخلايا العصبية في المسار nigrostriatal.
جانب آخر حاسم من نموذج الفأر هذا لمرض باركنسون هو النقطة الزمنية المطلوبة لتحليل عملية معينة بعد الجراحة. في هذا الصدد ، يظهر هذا العمل دراسة حركية للعمليات المختلفة التي ينطوي عليها علم الأمراض. نظرا لأن النقاط الزمنية الرئيسية تتغير مع جرعة الجينوم الفيروسي المعطاة لكل فأر ، فإن النمط المصلي ل AAV المستخدم ، أو حتى مع دفعة AAV المستخدمة ، تم إجراء تحليل استجابة الجرعة لكمية AAV-αSyn المطلوبة للحث على فقدان كبير في الخلايا العصبية TH+ والإعاقة الحركية لأول مرة. وقد أظهرت الدراسات السابقة ضعفا حركيا كبيرا وفقدان الخلايا العصبية TH+ في المسار nigrostriatal بعد 12 أسبوعا من حقن AAV-αSyn في الفئران بجرعات تتراوح بين 6 × 108-3 × 10 10 الجينوم الفيروسي لكل فأر 16,17,30,31. وفقا لذلك ، تراوحت جرعة AAV-hαSyn المستخدمة للحث على تعبير hαSyn في المسار nigrostriatal ، وفقدان الخلايا العصبية TH + ، والضعف الحركي في الفئران من 1 × 108-1 × 1010 الجينوم الفيروسي لكل فأر. علاوة على ذلك ، للسيطرة على أن فقدان الخلايا العصبية TH + والضعف الحركي قد تم إجراؤه بسبب الإفراط في التعبير عن hαSyn في SN وليس عن طريق عدوى AAV للخلايا العصبية في SN ، تم تضمين مجموعات التحكم التي تم فيها تسليم ترميز AAV لجين مراسل (AAV-eGFP) من جانب واحد في SN من الفئران وتم تحديد التنكس العصبي والإعاقة الحركية. أظهرت النتائج أنه بعد 12 أسبوعا من الجراحة المجسمة ، كان 1 ×10 10 جينومات فيروسية لكل فأر جرعة مناسبة من AAV5-hαSyn ، حيث أظهرت الفئران التي تتلقى هذا الحمل الفيروسي hαSyn كبيرا في المسار nigrostriatal (الشكل 2 والشكل 3) ، وفقدان الخلايا العصبية TH+ (الشكل 4) ، وضعف الحركة (الشكل 5). وعلى النقيض من ذلك، لم تكن الجرعات المنخفضة من AAV5-hαSyn (1 × 108 جينومات فيروسية لكل فأر و 1 × 109 جينومات فيروسية لكل ماوس) قوية بما يكفي للوصول إلى تغييرات كبيرة في جميع هذه المعلمات معا (الأشكال 2-4). تجدر الإشارة إلى أن إعطاء AAV-GFP عند 1 × 10 10 جينوم فيروسي لكل فأر أدى إلى انخفاض (~20٪) ، ولكن درجة كبيرة من فقدان الخلايا العصبية TH+ من الخلايا العصبية الدوبامينية النيجيرية (الشكل 4A ، B). تتفق هذه النتيجة مع الملاحظات السابقة باستخدام هذا النموذج41 وربما تكون نتيجة لانخفاض مستوى الالتهاب العصبي الناجم عن إعطاء ناقلات AAV في SN. ومع ذلك ، كان مدى فقدان الخلايا العصبية TH+ أعلى بكثير في الفئران التي تتلقى AAV5-hαSyn مقارنة بتلك التي تتلقى نفس الجرعة من AAV-GFP (الشكل 4C). تجدر الإشارة إلى أن حركية تعبير hαSyn لا تعتمد فقط على كفاءة النقل ولكن أيضا على مدى انتشار AAV39. نظرا لأن انتشار AAV يعتمد على النمط المصلي AAV ، فقد تختلف النقاط الزمنية الرئيسية الدقيقة في هذا النموذج الحيواني عند استخدام نمط مصلي AAV آخر مختلف عن AAV5.
بعد ذلك ، تم إجراء تحليل حركي باستخدام 1 × 1010 جينوم فيروسي لكل فأر لتحديد النقاط الزمنية الرئيسية في نموذج الماوس هذا. نظرا لأن الأدلة الحالية أظهرت بعض الأعراض المبكرة التي تظهر قبل الإعاقة الحركية ، والتي من شأنها أن تسمح بالتشخيص المبكر لمرض باركنسون43,44 ، سعت هذه التجارب إلى العثور على النقطة الزمنية التي كان فيها تعبير hαSyn واضحا بالفعل ولكن في غياب ضعف حركي. تظهر النتائج أن بداية تعبير hαSyn في SN كانت في 5 أسابيع بعد التسليم المجسمة ل AAV-hαSyn (الشكل 6). تشكل هذه النقطة الزمنية نقطة زمنية مثيرة للاهتمام لبدء إدارة العلاجات المصممة خصيصا لوقف العمليات العصبية الالتهابية والتنكسية العصبية. كانت النقاط الزمنية الرئيسية الأخرى المحددة هنا هي أوقات الذروة لحدثين حاسمين مرتبطين بعملية الالتهاب العصبي: الوقت الذي تصل فيه الخلايا الدبقية الصغيرة إلى أقصى درجة من التنشيط ووقت أقصى تسرب للخلايا التائية إلى SN. أظهرت النتائج منحنى مع اتجاه يصل إلى موجتين من التنشيط الدبقي الصغير الأقصى ، الأولى في 10 أسابيع بعد الجراحة والثانية في 15 أسبوعا بعد الجراحة (الشكل 7). أظهر التحليل الحركي لتسلل الخلايا التائية وقت الذروة لتسلل Treg إلى SN في 11 أسبوعا بعد الجراحة المجسمة (الشكل 8). والمثير للدهشة أنه لم يتم الكشف عن أي خلايا T مستجيبة (CD4 + Foxp3-) تتسلل إلى SN خلال الإطار الزمني الذي تم تحليله (الأسابيع 8-13 بعد الجراحة). إجمالا، تشير هذه النتائج إلى إطار زمني مناسب للبدء في إدارة العلاجات الموجهة نحو وقف عملية التهاب الأعصاب وتخفيف تسلل الخلايا التائية إلى SN باستخدام هذا النموذج قبل السريري، والذي يتراوح بين الأسبوع 5 بعد الجراحة (بداية الإفراط في التعبير عن hαsyn) والأسبوع 10 بعد الجراحة (الموجة الأولى من التهاب الأعصاب وتسلل الخلايا التائية) (الشكل 9).
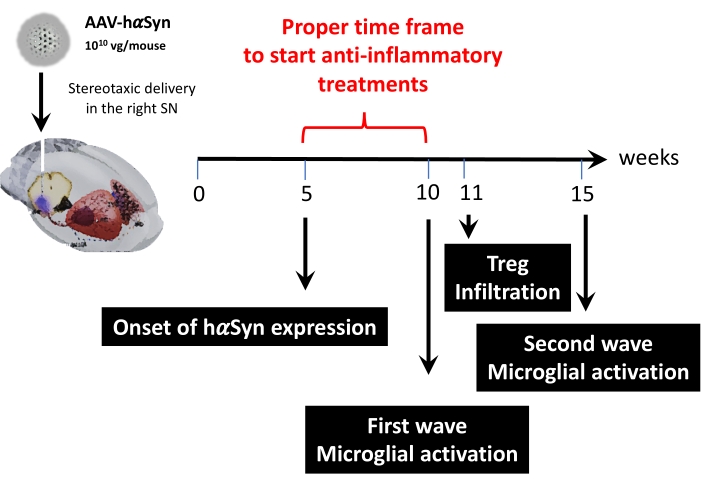
الشكل 9: ملخص للنقاط الزمنية الرئيسية التي تم العثور عليها لهذا النموذج الحيواني. يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
