Musemodellen for neurodegeneration, der analyseres her, kan hjælpe med at studere mange kritiske aspekter involveret i patofysiologien af Parkinsons sygdom, herunder de mekanismer, der er involveret i αSynpatologi og mikroglial aktivering, inddragelsen af det perifere immunsystem i reguleringen af neuroinflammation og mekanismerne for neurodegeneration. Blandt de mekanismer, der er involveret i αSyn-patologi, er de subcellulære mekanismer forbundet med mitokondriel, lysosomal eller proteasomal dysfunktion i nærvær af en overdreven belastning af αSyn i de dopaminerge neuroner i SN2. Det er vigtigt at overveje, at ud over hαSyn-ekspressionen induceret af AAV-medieret transduktion bidrager den endogene mus αSyn også til belastningen af det samlede αSyn-udtryk. Transgene mus, der overudtrykker mus αSyn, udvikler lignende synucleinpatologi, neuropatologi og motorisk svækkelse som disse musemodeller baseret på overekspression af hαSyn32. Med hensyn til mikroglial aktivering kan den nuværende musemodel bruges til at undersøge, hvordan forskellige molekylære og cellulære aktører såsom cytokiner, neurotransmittere, astrocytter, neuroner, blod-hjerne-barrieren og T-celler kan regulere erhvervelsen af proinflammatoriske eller antiinflammatoriske funktionelle fænotyper 8,10,11 . Denne model udgør også et vigtigt redskab til at studere det perifere immunsystems rolle, herunder ikke kun T-celler, men også makrofager, monocytter og neutrofiler, på processerne for neuroinflammation og neurodegeneration af nigrale neuroner 11,33,34. Endelig repræsenterer denne musemodel også et værdifuldt system til at studere de cellulære og molekylære mekanismer for neurodegeneration in vivo, herunder dem, der induceres af interne cellulære processer, såsom oxidativ stress, energiunderskud og beskadigede organeller 2, eller dem, der udøves af eksterne aktører, såsom neurotoksiske faktorer produceret af mikrogliaceller, astrocytter og cytotoksiske T-celler8, 28,29,35.
En begrænsning af denne musemodel er undersøgelsen af, hvordan den patologiske aggregering af αSyn på ekstra-cerebrale steder kan udgøre de indledende faser i udviklingen af Parkinsons sygdom36. I denne henseende er der voksende beviser, der tyder på, at før neurodegenerationen af nigralneuroner og motorisk svækkelse begynder αSynpatologi i tarmslimhinden og det olfaktoriske epitel36 og sandsynligvis også det αSyn-specifikke T-cellerespons12. Bagefter ville αSyn-aggregater migrere gennem vagusnerven til hjernestammen, hvilket udløste neuroinflammation og neurodegeneration af dopaminerge neuroner12. Selvom AAV-hαSyn-modellen rekapitulerer de fleste aspekter af Parkinsons sygdom, er der ingen tydelig involvering af den patologiske aggregering af αSyn på ekstra-cerebrale steder i denne model. En alternativ model, der involverer hαSyn-patologi, der er egnet til at studere disse aspekter af Parkinsons sygdom, kan være transgene mus, der overudtrykker hαSyn under kontrol af Thy1-promotoren, Thy1-SNCA-modellen 37, hvor sygdomsudvikling er afhængig af tarmmikrobiotaen og involverer en tydelig gastrointestinal svækkelse38.
Selvom det er nyttigt til undersøgelsen af de forskellige processer, der er forbundet med patofysiologien af Parkinsons sygdom, involverer den nuværende musemodel kritiske trin, der skal kontrolleres minutiøst, herunder korrekt levering af de virale vektorer i de tilsvarende rumlige koordinater, den selektive ekspression af hαSyn i neuroner (hvilket afhænger af AAV-serotypen og vektorkonstruktionen), og den korrekte AAV-dosis og timing, før du analyserer Parkinsons fænotype. Analysen af den korrekte levering af de virale vektorer i SN er nødvendig, da brugen af de korrekte rumlige koordinater for SN muligvis ikke er nok, når nålen ikke er helt lige, hvilket undertiden er umærkeligt for det menneskelige øje. Desuden afhænger diffusionen af AAV-vektorerne af AAV-serotype39. Af disse grunde er det nødvendigt at udføre periodiske kvalitetskontroller, der kontrollerer den korrekte levering og diffusion af de injicerede AAV-GFP-vektorer efter observation af GFP i hjerneskiver, der indeholder SN-området.
Med hensyn til den selektive ekspression af hαSyn i neuroner kunne ekspressionen af hαSyn i princippet konstrueres til at blive kontrolleret af en promotor, der er selektiv for neuroner eller, endnu mere præcis, selektiv for dopaminerge neuroner, såsom brugen af TH-promotoren i AAV-vektorer til at inducere den selektive ekspression af gener i dopaminerge neuroner40 . Denne strategi virker imidlertid ikke, når det, der søges, er overekspression af genet af interesse. Af denne grund er det i den nuværende model vigtigt at anvende en stærk promotor (en promotor, der inducerer høj ekspression af downstream-genet) og AAV-serotyper med neuronal tropisme. I denne undersøgelse blev CBA-promotoren brugt som en stærk promotor til at inducere overekspression af hαSyn, og AAV5-serotypen blev anvendt til den virale vektor. Denne serotype er tidligere blevet brugt til at transducere muse- ogrotteneuroner 41,42. Her viste resultaterne, at 12 uger efter leveringen af AAV5-GFP i musens SN var den grønne fluorescens selektivt til stede på den ipsilaterale side af både SN og striatum (figur 1), hvilket indikerer den effektive transduktion af neuroner i den nigrostriatale vej.
Et andet kritisk aspekt af denne musemodel af Parkinsons sygdom er det tidspunkt, der kræves for at analysere en bestemt proces efter operationen. I denne henseende viser dette arbejde en kinetisk undersøgelse af forskellige processer involveret i patologien. Da vigtige tidspunkter ændrer sig med den dosis af virale genomer, der gives pr. mus, blev serotypen af AAV anvendt, eller endda med det anvendte parti AAV, først udført en dosis-respons-analyse af mængden af AAV-αSyn, der kræves for at inducere et signifikant tab af TH + neuroner og motorisk svækkelse. Tidligere undersøgelser har vist signifikant motorisk svækkelse og et tab af TH+ neuroner i den nigrostriatale vej efter 12 ugers AAV-αSyn-injektioner i mus i doser fra 6 x 108-3 x 1010 virale genomer pr. mus 16,17,30,31. Følgelig varierede dosis af AAV-hαSyn, der blev anvendt til at inducere hαSyn-ekspressionen i den nigrostriatale vej, tabet af TH+ neuroner og motorisk svækkelse hos mus fra 1 x 108-1 x 1010 virale genomer pr. Mus. For at kontrollere, at tabet af TH + neuroner og motorisk svækkelse blev induceret af overekspression af hαSyn i SN og ikke ved AAV-infektion af neuroner i SN, blev kontrolgrupper inkluderet, hvor AAV-kodning for et reportergen (AAV-eGFP) blev leveret ensidigt i SN af mus, og neurodegeneration og motorisk svækkelse blev bestemt. Resultaterne viste, at 1 x 1010 virale genomer pr. mus 12 uger efter stereotaksisk kirurgi var en korrekt dosis AAV5-hαSyn, da mus, der modtog denne virale belastning, viste signifikant hαSyn i den nigrostriatale vej (figur 2 og figur 3), tab af TH + neuroner (figur 4) og motorisk svækkelse (figur 5). I modsætning hertil var lavere doser af AAV5-hαSyn (1 x 108 virale genomer pr. mus og 1 x 109 virale genomer pr. mus) ikke stærke nok til at nå signifikante ændringer i alle disse parametre tilsammen (figur 2-4). Bemærk, at administrationen af AAV-GFP ved 1 x 1010 virale genomer pr. Mus inducerede en lav (~ 20%), men signifikant grad af tab af TH + neuroner af nigral dopaminerge neuroner (figur 4A, B). Dette resultat stemmer overens med tidligere observationer ved hjælp af denne model41 og er sandsynligvis konsekvensen af et lavt niveau af neuroinflammation induceret af administrationen af AAV-vektorer i SN. Ikke desto mindre var omfanget af tab af TH+-neuroner signifikant højere hos mus, der fik AAV5-hαSyn sammenlignet med dem, der fik samme dosis AAV-GFP (figur 4C). Det bemærkes, at kinetikken af hαSyn-ekspression ikke kun afhænger af effektiviteten af transduktion, men også af omfanget af AAV-diffusion39. Da AAV-diffusion afhænger af AAV-serotypen, kan de præcise nøgletidspunkter i denne dyremodel variere, når der anvendes en anden AAV-serotype, der er forskellig fra AAV5.
Derefter blev der udført en kinetisk analyse ved hjælp af 1 x 1010 virale genomer pr. Mus for at bestemme vigtige tidspunkter i denne musemodel. Da de nuværende beviser har vist nogle tidlige symptomer, der vises før motorisk svækkelse, hvilket ville muliggøre tidlig diagnose af Parkinsons sygdom43,44, forsøgte disse eksperimenter at finde det tidspunkt, hvor hαSyn-ekspression allerede var tydelig, men i mangel af motorisk svækkelse. Resultaterne viser, at starten på hαSyn-ekspression i SN var 5 uger efter den stereotaksiske levering af AAV-hαSyn (figur 6). Dette tidspunkt udgør et interessant tidsmæssigt punkt at begynde at administrere terapier, der er skræddersyet til at stoppe de neuroinflammatoriske og neurodegenerative processer. Andre vigtige tidspunkter, der blev bestemt her, var spidsbelastningstiderne for to kritiske hændelser forbundet med neuroinflammationsprocessen: det tidspunkt, hvor mikroglia når den maksimale grad af aktivering og tidspunktet for maksimal T-celleinfiltration i SN. Resultaterne viste en kurve med en tendens, der nåede to bølger af maksimal mikroglial aktivering, den første 10 uger efter operationen og den anden 15 uger efter operationen (figur 7). Den kinetiske analyse af T-celleinfiltration viste spidsbelastningstiden for Treg-infiltration i SN 11 uger efter stereotaksisk kirurgi (figur 8). Overraskende nok blev der ikke påvist nogen effektor T-celler (CD4 + Foxp3-), der infiltrerede SN i løbet af den analyserede tidsramme (uge 8-13 efter operationen). Alt i alt tyder disse resultater på en passende tidsramme for at begynde at administrere terapier rettet mod at stoppe processen med neuroinflammation og dæmpe T-celleinfiltration i SN ved hjælp af denne prækliniske model, som varierer mellem uge 5 efter operationen (begyndelsen af hαSyn overekspression) og uge 10 efter operationen (den første bølge af neuroinflammation og T-celleinfiltration) (figur 9).
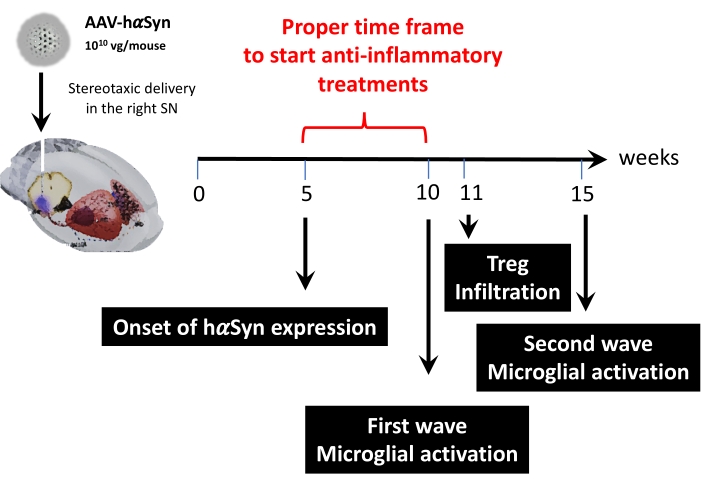
Figur 9: Oversigt over de vigtigste tidspunkter, der er fundet for denne dyremodel. Klik her for at se en større version af denne figur.
