Musmodellen för neurodegeneration som analyseras här kan hjälpa till att studera många kritiska aspekter som är involverade i patofysiologin vid Parkinsons sjukdom, inklusive mekanismerna som är involverade i αSyn-patologi och mikroglial aktivering, involveringen av det perifera immunsystemet i regleringen av neuroinflammation och mekanismerna för neurodegeneration. Bland de mekanismer som är involverade i αSyn-patologi är de subcellulära mekanismerna associerade med mitokondriell, lysosomal eller proteasomal dysfunktion i närvaro av en överdriven belastning av αSyn i de dopaminerga neuronerna iSN2. Det är viktigt att tänka på att förutom hαSyn-uttrycket inducerat av AAV-medierad transduktion bidrar den endogena musen αSyn också till belastningen av totalt aSyn-uttryck. Transgena möss som överuttrycker mus αSyn utvecklar liknande synukleinpatologi, neuropatologi och motorisk försämring som dessa musmodeller baserat på överuttrycket av hαSyn32. När det gäller mikroglial aktivering kan den nuvarande musmodellen användas för att studera hur olika molekylära och cellulära spelare såsom cytokiner, neurotransmittorer, astrocyter, neuroner, blod-hjärnbarriären och T-celler kan reglera förvärvet av proinflammatoriska eller antiinflammatoriska funktionella fenotyper 8,10,11 . Denna modell utgör också ett viktigt verktyg för att studera det perifera immunsystemets roll, inklusive inte bara T-celler utan även makrofager, monocyter och neutrofiler, på processerna för neuroinflammation och neurodegenerering av nigralneuroner 11,33,34. Slutligen representerar denna musmodell också ett värdefullt system för att studera de cellulära och molekylära mekanismerna för neurodegeneration in vivo, inklusive de som induceras av interna cellulära processer, såsom oxidativ stress, energiunderskott och skadade organeller 2, eller de som utövas av externa aktörer, såsom neurotoxiska faktorer som produceras av mikrogliaceller, astrocyter och cytotoxiska T-celler8, 28,29,35.
En begränsning av denna musmodell är studien av hur den patologiska aggregeringen av αSyn på extra-cerebrala platser kan utgöra de inledande stadierna i utvecklingen av Parkinsons sjukdom36. I detta avseende finns det växande bevis som tyder på att αSyn-patologin börjar i tarmslemhinnan och luktepitelet36 och förmodligen det αSyn-specifika T-cellsvaret12 före neurodegenerationen av nigralneuroner och motorisk försämring. Efteråt skulle αSyn-aggregat migrera genom vagusnerven till hjärnstammen, vilket utlöser neuroinflammation och neurodegeneration av dopaminerga neuroner12. Även om AAV-hαSyn-modellen rekapitulerar de flesta aspekter av Parkinsons sjukdom, finns det ingen uppenbar inblandning av den patologiska aggregeringen av αSyn på extra-cerebrala platser i denna modell. En alternativ modell som involverar hαSyn-patologi som är lämplig för att studera dessa aspekter av Parkinsons sjukdom kan vara transgena möss som överuttrycker hαSyn under kontroll av Thy1-promotorn, Thy1-SNCA-modellen 37, där sjukdomsutvecklingen är beroende av tarmmikrobiotan och innebär en uppenbar gastrointestinal försämring38.
Även om det är till hjälp för studien av de olika processerna som är associerade med patofysiologin vid Parkinsons sjukdom, innebär den nuvarande musmodellen kritiska steg som bör kontrolleras noggrant, inklusive korrekt leverans av virusvektorerna i motsvarande rumsliga koordinater, det selektiva uttrycket av hαSyn i neuroner (vilket beror på AAV-serotypen och vektorkonstruktionen), och rätt AAV-dos och timing innan du analyserar den parkinsoniska fenotypen. Analysen av korrekt leverans av virusvektorerna i SN är nödvändig, eftersom användningen av de korrekta rumsliga koordinaterna för SN kanske inte räcker när nålen inte är helt rak, vilket ibland är omärkligt för det mänskliga ögat. Dessutom beror diffusionen av AAV-vektorerna på AAV-serotyp39. Av dessa skäl är det nödvändigt att utföra regelbundna kvalitetskontroller för att kontrollera korrekt leverans och diffusion av de injicerade AAV-GFP-vektorerna efter observation av GFP i hjärnskivor som innehåller SN-området.
När det gäller det selektiva uttrycket av hαSyn i neuroner kan uttrycket av hαSyn i princip konstrueras för att styras av en promotor selektiv för neuroner eller, ännu mer exakt, selektiv för dopaminerga neuroner, såsom användningen av TH-promotorn i AAV-vektorer för att inducera det selektiva uttrycket av gener i dopaminerga neuroner40 . Denna strategi fungerar dock inte när det som eftersträvas är överuttryck av genen av intresse. Av denna anledning är det i den nuvarande modellen viktigt att använda en stark promotor (en promotor som inducerar högt uttryck av nedströmsgenen) och AAV-serotyper med neuronal tropism. I denna studie användes CBA-promotorn som en stark promotor för att inducera överuttryck av hαSyn, och AAV5-serotypen användes för virusvektorn. Denna serotyp har använts tidigare för att transducera mus- och råttneuroner41,42. Här visade resultaten att 12 veckor efter leveransen av AAV5-GFP i SN hos möss var den gröna fluorescensen selektivt närvarande på ipsilateralsidan av både SN och striatum (Figur 1), vilket indikerar effektiv transduktion av neuroner i den nigrostriatala vägen.
En annan kritisk aspekt av denna musmodell av Parkinsons sjukdom är den tidpunkt som krävs för att analysera en viss process efter operationen. I detta avseende visar detta arbete en kinetisk studie av olika processer som är involverade i patologin. Eftersom viktiga tidpunkter förändras med dosen av virala genom som ges per mus, den serotyp av AAV som används, eller till och med med den sats AAV som används, utfördes först en dos-responsanalys av mängden AAV-αSyn som krävs för att inducera en signifikant förlust av TH + -neuroner och motorisk försämring. Tidigare studier har visat signifikant motorisk försämring och en förlust av TH+ neuroner i nigrostriatalvägen efter 12 veckors AAV-αSyn-injektioner hos möss i doser från 6 x 108–3 x 1010 virala genom per mus 16,17,30,31. Följaktligen varierade dosen av AAV-hαSyn som användes för att inducera hαSyn-uttrycket i den nigrostriatala vägen, förlusten av TH + -neuroner och motorisk försämring hos möss från 1 x 108–1 x 1010 virala genom per mus. För att kontrollera att förlusten av TH + -neuroner och motorisk försämring inducerades av överuttrycket av hαSyn i SN och inte av AAV-infektion av neuroner i SN, inkluderades kontrollgrupper där AAV-kodning för en reportergen (AAV-eGFP) levererades ensidigt i SN hos möss och neurodegeneration och motorisk försämring bestämdes. Resultaten visade att 1 x10 10 virusgenom per mus 12 veckor efter stereotaxisk kirurgi var en korrekt dos av AAV5-hαSyn, eftersom möss som fick denna virusbelastning visade signifikant hαSyn i den nigrostriatala vägen (figur 2 och figur 3), förlust av TH + -neuroner (figur 4) och motorisk försämring (figur 5). Däremot var lägre doser av AAV5-hαSyn (1 x 108 virala genom per mus och 1 x 109 virala genom per mus) inte tillräckligt starka för att nå signifikanta förändringar i alla dessa parametrar tillsammans (figur 2–4). Observera att administreringen av AAV-GFP vid 1 x 1010 virala genom per mus inducerade en låg (~ 20%), men signifikant grad av förlust av TH + -neuroner av nigral dopaminerga neuroner (Figur 4A, B). Detta resultat överensstämmer med tidigare observationer med denna modell41 och är förmodligen konsekvensen av en låg nivå av neuroinflammation inducerad av administrering av AAV-vektorer i SN. Ändå var omfattningen av förlusten av TH+ neuroner signifikant högre hos möss som fick AAV5-hαSyn jämfört med dem som fick samma dos av AAV-GFP (figur 4C). Observera att kinetiken för hαSyn-uttryck inte bara beror på transduktionens effektivitet utan också på omfattningen av AAV-diffusion39. Eftersom AAV-diffusion beror på AAV-serotypen kan de exakta nyckeltidpunkterna i denna djurmodell variera när man använder en annan AAV-serotyp som skiljer sig från AAV5.
Därefter genomfördes en kinetisk analys med 1 x 1010 virala genom per mus för att bestämma viktiga tidpunkter i denna musmodell. Eftersom nuvarande bevis har visat några tidiga symtom som uppträder före motorisk försämring, vilket skulle möjliggöra tidig diagnos av Parkinsons sjukdom43,44, försökte dessa experiment hitta den tidpunkt då hαSyn-uttrycket redan var uppenbart men i avsaknad av motorisk försämring. Resultaten visar att uppkomsten av hαSyn-uttryck i SN var 5 veckor efter stereotaxisk leverans av AAV-hαSyn (figur 6). Denna tidpunkt utgör en intressant tidspunkt för att börja administrera terapier skräddarsydda för att stoppa de neuroinflammatoriska och neurodegenerativa processerna. Andra viktiga tidpunkter som bestämdes här var topptiderna för två kritiska händelser associerade med neuroinflammationsprocessen: den tid då mikroglia når maximal aktiveringsgrad och tiden för maximal T-cellinfiltration i SN. Resultaten visade en kurva med en trend som nådde två vågor av maximal mikroglial aktivering, den första vid 10 veckor efter operationen och den andra vid 15 veckor efter operationen (figur 7). Den kinetiska analysen av T-cellsinfiltration visade topptiden för Treg-infiltration i SN vid 11 veckor efter stereotaxisk operation (figur 8). Överraskande nog upptäcktes inga effektor T-celler (CD4 + Foxp3-) som infiltrerade SN under den analyserade tidsramen (vecka 8–13 efter operationen). Sammantaget föreslår dessa resultat en lämplig tidsram för att börja administrera terapier inriktade på att stoppa processen med neuroinflammation och dämpa T-cellinfiltration i SN med hjälp av denna prekliniska modell, som sträcker sig mellan vecka 5 efter operationen (uppkomsten av hαSyn-överuttryck) och vecka 10 efter operationen (den första vågen av neuroinflammation och T-cellinfiltration) (Figur 9).
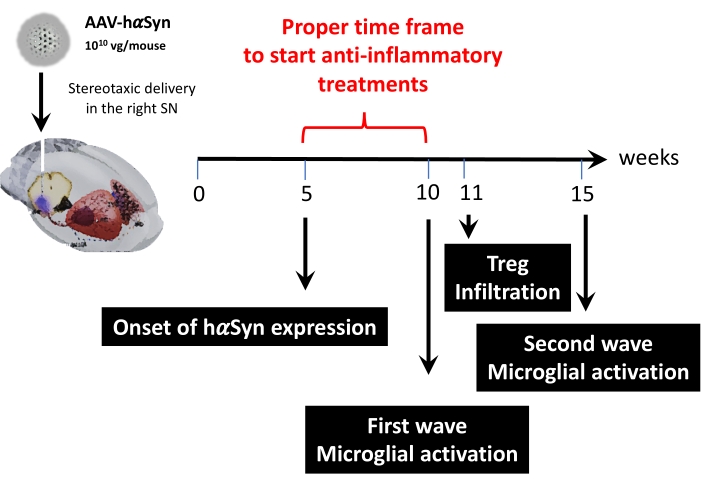
Figur 9: Sammanfattning av de viktigaste tidpunkterna som hittades för denna djurmodell.
