Durch die Abgabe und Entfernung von Proteinen zur und von der Plasmamembran steuert der Membranproteintransport in Neuronen die Plasmamembranausrüstung mit Rezeptoren sowie Ionenkanälen und reguliert dadurch die neuronale Funktion. Fehlregulationen oder Defekte im Proteintransport haben typischerweise schädliche Auswirkungen auf Zellen und führen zu neuronaler Degeneration. Beim Menschen kann dies neurodegenerative Erkrankungen wie Alzheimer und Parkinson oder Retinitis pigmentosa1 verursachen. Photorezeptoren im Facettenauge von Drosophila melanogaster sind zu einem In-vivo-Modellsystem für die Untersuchung des Membranproteintransportsgeworden 2. Dies liegt nicht nur an der genetischen Vielseitigkeit von Drosophila , die effektive genetische Screens ermöglicht, sondern auch daran, dass alle wesentlichen Bestandteile der lichtabsorbierenden Photorezeptormembran sehr detailliert charakterisiert sind und effiziente mikroskopische Techniken zur Verfügung stehen, die auf das Fliegenauge angewendet werden können. Diese Techniken stehen im Mittelpunkt dieses Artikels.
In Drosophila-Photorezeptorzellen bildet die apikale Plasmamembran einen dicht gepackten Stapel von Mikrovilli entlang einer Seite der Zelle, der als Rhabdomere bezeichnet wird. Die Rhabdomere der Photorezeptorzellen R1-6 sind in einem charakteristischen trapezförmigen Muster angeordnet, während die Photorezeptorzellen R7 und R8 ein einzelnes Rhabdomere in der Mitte diesesTrapezes 3 bilden. Der Membranproteintransport wird für einen regulierten Umsatz von rhabdomeralen Membranproteinen wie Rhodopsin und den lichtaktivierten Ionenkanälen TRP (transient receptor potential) und TRPL (TRP-like) benötigt, um die richtige Menge dieser Phototransduktionsproteine im Rhabdomere sicherzustellen. Photorezeptormembranproteine werden im endoplasmatischen Retikulum synthetisiert und über den Golgi-Apparat zum Rhabdomere transportiert. Nach der Aktivierung von Rhodopsin durch Licht kann ein Rhodopsinmolekül entweder durch Absorption eines zweiten Photons inaktiviert oder durch Clathrin-vermittelte Endozytose aus dem Rhabdomere entfernt werden. Endozytosiertes Rhodopsin wird entweder im Lysosom abgebaut oder in den Rhabdomerezurückgeführt 4,5. Der Ionenkanal TRPL wird nach Aktivierung der Phototransduktionskaskade ebenfalls internalisiert und durchläuft eine lichtabhängige Translokation zwischen dem Rhabdomere (wo er sich befindet, wenn Fliegen im Dunkeln gehalten werden) und einem ER-angereicherten Staufach im Zellkörper (zu dem er bei Beleuchtung innerhalb weniger Stunden transportiert wird)6,7,8,9,10 . Im Gegensatz zu endozytosiertem Rhodopsin werden nur geringe Mengen TRPL über den endolysosomalen Weg abgebaut, und der Großteil wird stattdessen intrazellulär gespeichert und bei dunkler Anpassung wieder in den Rhabdomer zurückgeführt6. TRPL kann somit zur Analyse des lichtgetriggerten Transports von Plasmamembranproteinen verwendet werden. Drosophila-Photorezeptorzellen werden auch zur Untersuchung der neuronalen Degeneration eingesetzt. Die Degeneration von Photorezeptorzellen wird häufig durch die Beurteilung der Struktur von Rhabdomeren bestimmt, die infolge degenerativer Prozesse zerfallen5.
Um die subzelluläre Lokalisation von TRPL und Rhodopsin in Photorezeptorzellen oder die Degeneration von Photorezeptorzellen zu untersuchen, wurden hier zwei Fluoreszenzmikroskopie-Methoden angewendet, die sich hinsichtlich Analysegeschwindigkeit und Auflösung unterscheiden. Eine sehr schnelle, nicht-invasive Methode, die für genetische Screens verwendet werden kann, jedoch mit einer begrenzten räumlichen Auflösung, ist der Nachweis der Fluoreszenz in der tiefen Pseudopupille (DPP). Das DPP ist ein optisches Phänomen von Arthropoden-Facettenaugen, dessen geometrischer Ursprung1971 von Franceschini und Kirschfeld ausführlich erklärt wurde 11. Kurz gesagt, auf mehreren optischen Ebenen unterhalb der Netzhautüberlagerung können Bilder von Rhabdomeren aus benachbarten Ommatidien beobachtet werden. Auf einer Fokusebene durch die Mitte der Augenkrümmung bilden diese überlagerten Projektionen ein Bild, das dem trapezförmigen Layout von Rhabdomeren in einem einzigen Ommatidium ähnelt, das nur um Größenordnungen größer ist. Dieses Phänomen kann auch unabhängig von der exogenen Expression von Fluoreszenzproteinen (z.B. TRPL::EGFP 8) beobachtet werden, was den Nachweis des DPP jedoch erleichtert (Abbildung 1A-A‘‘)12. Eine zweite nicht-invasive Methode ist die Wasserimmersionsmikroskopie, die auf der Abbildung fluoreszierend markierter Proteine beruht, nachdem der Dioptrienapparat der Augen optisch mit Wasser neutralisiert wurde (Abbildung 1B-C‘‘)12. Mit der Wasserimmersionsmethode kann die relative Menge an TRPL::eGFP in den Rhabdomeren oder im Zellkörper für einzelne Photorezeptorzellen quantitativ beurteilt werden. Darüber hinaus können nicht-translozierende fluoreszenzmarkierte Proteine verwendet werden, um die rhabdomerale Integrität zu bewerten und den zeitlichen Verlauf einer möglichen Degeneration quantitativ zu bestimmen, wie hier beschrieben.
Während Aufzeichnungen des DPP bei weitem die einfachste und schnellste dieser Methoden sind, ist die räumliche Auflösung der von ihnen generierten Daten begrenzt. Darüber hinaus gibt es zahlreiche Gründe, warum ein DPP fehlen kann, die für die DPP-Bildgebung selbst nicht unbedingt erkennbar sind. Da der DPP eine Summe mehrerer Ommatidien darstellt, gehen Informationen über einzelne Zellen verloren. Daher erfüllt die DPP-Bildgebung mit niedriger Auflösung eine wichtige Funktion beim Screening einer großen Anzahl von Fliegen, sollte jedoch im Allgemeinen von Aufnahmen mit höherer Auflösung mittels Wasserimmersionsmikroskopie gefolgt werden. Wasserimmersionsmikroskope erlauben Interpretationen über einzelne Zellen, Entwicklungsdefekte, Augenmorphologie, Proteinfehllokalisation oder Netzhautdegeneration sowie die Quantifizierung dieser Effekte. Dieses Protokoll beschreibt diese beiden Techniken im Detail.
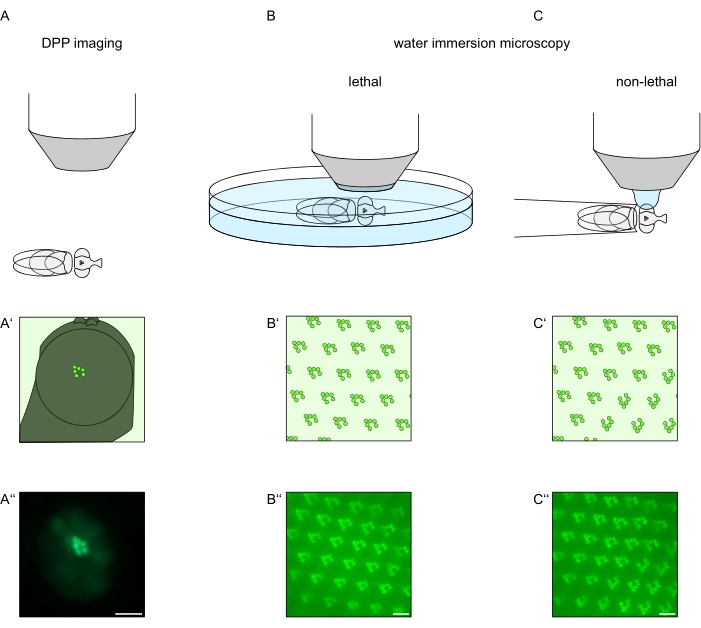
Abbildung 1: Übersicht über die in diesem Protokoll dargestellten Mikroskopievariationen für das Drosophila-Auge. Schematische Darstellungen und exemplarische Mikroaufnahmen der (A-A”) fluoreszierenden tiefen Pseudopupillenbildgebung (DPP), (B-B”) der tödlichen Wasserimmersionsmikroskopie von fluoreszierenden Rhabdomeren und (C-C‘‘) der nicht-tödlichen Wassertropfenmikroskopie von fluoreszierenden Rhabdomeren. Maßstabsleiste (A”): 100 μm. Maßstabsstäbe (B”–C‘‘): 10 μm Die Abbildung wurde gegenüber Referenz13 geändert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
