We show representative outcomes from Step 3 onwards. Figure 1 shows the intact explanted bowels, with fluorescent measurements overlaid. The stomach (purple) is laid along the same axis as the small intestine (orange), but we prefer moving the cecum (blue) to the side to prevent overlap with the large intestine (orange). As evidenced in the left panel, this is not always possible due to organ size. We cut the small bowel at ~200 mm to maximize the coverage of continuous segments, but this is not always possible due to mesentery limiting the ability to unfold the intestines, and our preference not to cut through structures like fluorescent pellets or the cecum. The red intensity in the left panel of Figure 1 and the green intensities of the middle and right panels are sorted into different sized bins to generate Figure 3. Only the intensities within the orange ROIs (small bowel) are extracted for analysis. The full length of the small bowel (every orange ROI in each panel) is extracted for analysis. Orange ROIs are the same width, as displayed in ImageJ (Figure 2). They are stitched together by placing all intensities in sequence prior to binning. Figure 3 shows the average fluorescent trace ± SEM per cohort; see figure legend for specifics.
The last two figures demonstrate the results of the analyses: geometric center and power spectrum. The geometric center measures the average location of the fluorescent signals in Figure 3, weighing the average by the signal strength of each bin. Hence, higher peaks in the trace pull the average closer to the position of that peak. The geometric center fails to completely characterize the spatial distribution of intraluminal contents. For example, the same geometric center can be obtained from a bolus concentrated at a single point and a spread-out signal centered around the same point. This limitation of the geometric center measurement is evident in the comparison between liquids and larger beads (Figure 3 and Figure 4). The majority of the liquid fluorescent trace is centered around two early peaks, whereas the larger beads show multiple peaks along the full length of the small bowel (Figure 3, left and right panels). The fluorescent trace of the larger beads is more distributed, but it averages out to a similar point as that of the liquid, regardless of binning granularity, which highlights the limitation of solely focusing on the geometric center (Figure 4). To account for the distributive nature of small bowel contractions, we have incorporated a power spectral analysis into the protocol. The power spectrum analysis works by deconstructing the fluorescence distribution in space into multiple sinusoid curves of different spatial frequencies. Each spatial frequency reflects the distance between two randomly selected points in the fluorescence trace. Some of these distances occur more frequently than others, so each is attributed a strength (power). The power correlates with how frequently two random points are separated by that distance. By plotting the power spectrum (Figure 5), we can demonstrate how many spatial frequencies contribute to the measured signal (spread of the spectrum) and compare the relative strength of those contributions (height of the spectrum). This allows us to quantitatively describe the spread of fluorescence along the GI tract.
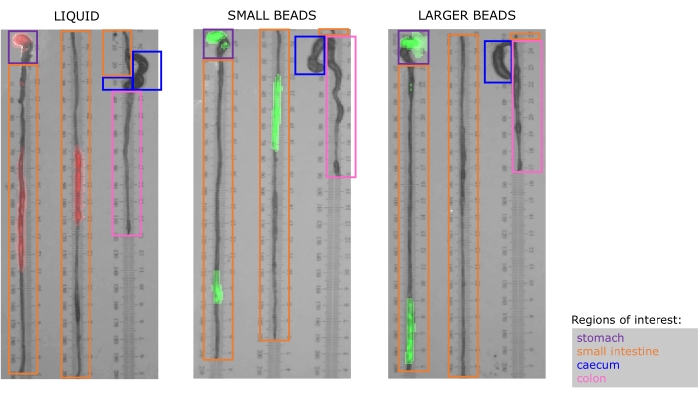
Figure 1. The dissected bowels are explanted and placed on laminated ruler paper prior to photography and fluorescence measurement. Fluorescence intensity is overlayed and pseudocolored to match the native fluorescence of gavaged material. Left: liquid rhodamine isothiocyanate (RITC) fluoresces in the red spectrum; Middle: small (diameter: 75-90 µm), and Right: larger (diameter: 180-212 µm) beads both fluoresce in the green spectrum. Purple, orange, blue, and pink boxes surround regions of interest (ROIs) in the stomach, small intestine, cecum, and colon in each specimen, respectively. The width of each ROI is kept consistent across rows to avoid confusion when calculating the raw fluorescence intensity during downstream analysis. Please click here to view a larger version of this figure.
Figure 2. Location of the region of interest width (in pixels) in the ImageJ toolbar (yellow underline). To reiterate, the width is displayed in pixels only if the scale has been removed (Step 5.10). Please click here to view a larger version of this figure.
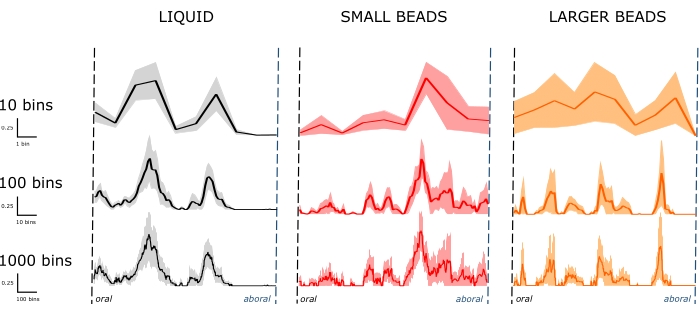
Figure 3. Fluorescence traces along the length of the small bowel. Line denotes the average fluorescence at a given bin. The shaded area denotes the standard error of the mean (SEM). The distribution of fluorescent material varies according to the material properties of the intraluminal contents (columns). Left: liquid (n = 7 mice) distribution across a few small bowel segments 30 min after gavage. Middle and right: small (n = 6 mice) and larger (n = 5 mice) beads distribute more broadly along the small bowel 30 min after gavage. Increasing the number of bins used to sort the same raw data sets reveals granular trace features unresolvable with fewer bins (rows). Smaller bins decrease measurement uncertainty, increase spatial resolution and better reflect the distributive component of small bowel contractions. Please click here to view a larger version of this figure.
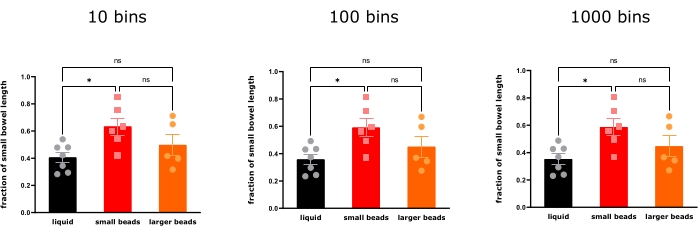
Figure 4. The geometric center of the fluorescence distribution in small bowel 30 min after gavage with liquid RITC, small beads (75-90 µm) and larger beads (180-212 µm) (mean ± SEM, n = 7, n = 6, n = 5, one-way ANOVA with Bonferroni correction). Bin size does not affect the transit interpretation that small beads are transported more distally than liquids 30 min after gavage, but the larger beads distribute so widely that there are no significant differences in their geometric center compared to both liquids and small beads (*P < 0.05. ns = not statistically significant). Please click here to view a larger version of this figure.
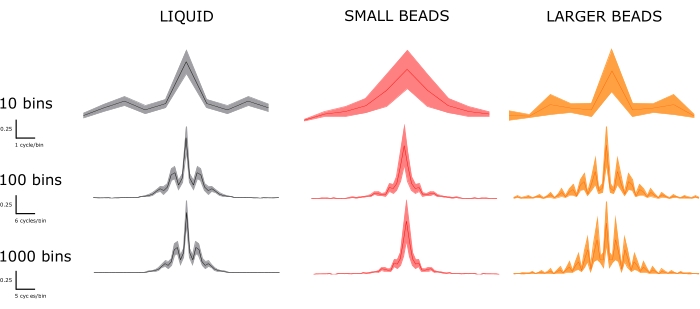
Figure 5. Power spectra are categorized by material property (columns) and bin size (rows). The improvements in spatial resolution and smaller binning are evident. Top row: few differences can be distinguished in the widely binned spectra. Bottom row: as bin sizes shrink, we can appreciate significant dominant frequencies present in the larger beads' spectrum but not in that of the small beads. Some, but not all, of those additional dominant frequencies, are present in the liquid spectrum. Using the bottom row spectra, we can compare to the fluorescent traces in Figure 3. In Figure 3, the liquid fluorescent trace displays two prominent peaks, which correlates with a few dominant peaks in the power spectrum. The small beads trace in Figure 3 displays a single dominant peak, which correlates with a single dominant peak in the power spectrum. The larger beads trace in Figure 3 displays more dominant peaks. Accordingly, the power spectrum shows a greater number of dominant frequencies. Please click here to view a larger version of this figure.
