Die Lipidinfiltration der Skelettmuskulatur, bekannt als Myosteatose, nimmt mit Fettleibigkeit und Alterung zu. Die Myosteatose korreliert negativ mit Muskelmasse und Kraft und mit der Insulinsensitivität1. Darüber hinaus deuten neuere Studien darauf hin, dass der Grad der Myosteatose als prognostischer Faktor für andere Erkrankungen wie Herz-Kreislauf-Erkrankungen2, nicht-alkoholische Fettlebererkrankungen3 oder Krebs4 verwendet werden könnte. Lipide können sich in der Skelettmuskulatur zwischen Muskelfasern als extramyozelluläre Lipide oder innerhalb der Fasern, als intramyozelluläre Lipide (IMCLs), ansammeln. IMCLs werden überwiegend als Triglyceride in Lipidtröpfchen (LDs) gespeichert, die als Stoffwechselbrennstoff bei körperlicher Betätigung verwendet werden 5,6. Wenn jedoch das Lipidangebot die Nachfrage übersteigt oder wenn Mitochondrien dysfunktional werden, werden IMCLs an der Muskelinsulinresistenz beteiligt, wie sie bei metabolisch ungesunden, fettleibigen Personen und bei Typ-2-Diabetes-Patientenbeobachtet wird 7. Interessanterweise haben Ausdauersportler ähnliche, wenn nicht sogar höhere IMCLs-Spiegel wie bei adipösen Patienten mit Typ-2-Diabetes mellitus, während sie gleichzeitig eine hohe Insulinsensitivität beibehalten. Dieses Phänomen wird als “Athletenparadoxon”8,9 beschrieben und wird durch eine nuanciertere Bewertung der Muskel-LDs in Bezug auf ihre Größe, Dichte, Lokalisation, Dynamik und Zusammensetzung der Lipidspezies erklärt.
Erstens korreliert die LD-Größe umgekehrt mit der Insulinsensitivität und der körperlichen Fitness10,11. Tatsächlich weisen kleinere LDs eine relativ größere Oberfläche für die Lipasewirkung auf und haben daher möglicherweise eine größere Fähigkeit, Lipide zu mobilisieren12. Zweitens spielt die LD-Dichte (Anzahl/Oberfläche) eine umstrittene Rolle bei der Insulinwirkung 8,10; Dennoch scheint es bei Athleten erhöht zu sein. Drittens ist die subzelluläre Lokalisation von LDs wichtig, da LDs, die sich direkt unter der Oberflächenmembran (subsarkolemmal oder peripher) befinden, eine schädlichere Wirkung auf die Insulinsensitivität ausüben als zentrale 8,9,13. Letztere liefern Brennstoff für zentrale Mitochondrien, die eine größere Atmungsaktivität haben und spezialisierter sind, um den hohen Energiebedarf zu decken, der für die Kontraktionerforderlich ist 14. Im Gegensatz dazu liefern periphere LDs subsarkolämale Mitochondrien, die an membranbezogenen Prozessen beteiligt sind8. Schließlich können über Triglyceride hinaus spezifische komplexe Lipide im Muskel schädlicher sein als andere. Zum Beispiel können sich Diacylglycerin, langkettiges Acyl-CoA und Ceramide im Muskel ansammeln, wenn die Triglycerid-Umsatzrate niedrig ist, wodurch die Insulinsignalisierung 9,15 beeinträchtigt wird. Um auf das “Athletenparadoxon” zurückzukommen: Ausdauersportler haben eine hohe Anzahl kleinerer zentraler LDs mit erhöhten Fluktuationsraten in Typ I (oxidativen) Fasern, während adipöse und diabetische Patienten größere periphere LDs mit niedrigen Fluktuationsraten in Typ II (glykolytischen) Fasern haben 8,15,16. Zusätzlich zu ihrer Rolle bei der Energiespeicherung und -freisetzung könnten LDs über abgeleitete Fettsäuren (FA) und ein Mantelprotein (Perilipin 5) auch als kritische Akteure fungieren, die an der transkriptionellen Regulation der FA-Oxidation und der mitochondrialen Biogenese beteiligt sind8. Aufgrund ihrer entscheidenden Implikationen in der Physiologie und Pathophysiologie sind eingehende Studien über die Dynamik und Funktionen von LDs gerechtfertigt.
Obwohl es mehrere Techniken gibt, um IMCLs zu untersuchen, sind sie nicht alle geeignet, LD-Größe, -Dichte und -Verteilung auf faserspezifische Weise genau zu quantifizieren. Zum Beispiel bietet die Bewertung von IMCLs durch Magnetresonanzspektroskopie zwar nicht-invasiv, bietet aber eine Auflösung, die nicht ausreicht, um die Größe und genaue Position von LDs innerhalb der Faser zu untersuchen, und sie ist nicht fasertypspezifisch17,18. Ebenso können biochemische Techniken, die an Ganzmuskelhomogenaten19 durchgeführt werden, die Lage und Größe von Lipiden nicht beurteilen. Folglich ist die am besten geeignete Methode zur Analyse der LD-Morphologie und -Ortung die quantitative elektronische Transmissionsmikroskopie13, aber diese Technik ist teuer und zeitaufwendig. Daher hat sich die konfokale Fluoreszenzbildgebung auf Präparaten mit Farbstoffen wie Oil Red O (ORO)20,21, Monodansylpentan (MDH)22 oder Bodipy23,24,25 als das beste Werkzeug für diese Studien herausgestellt.
Hier wird ein vollständiges Protokoll beschrieben, einschließlich Gewebeentnahme und -verarbeitung, Bodipy-Färbung und konfokaler Bildaufnahme und -analyse zur Quantifizierung der LD-Größe, -Anzahl und -Lokalisation in Mausmuskel-Kryosektionen. Da IMCLs nicht gleichmäßig auf oxidative und glykolytische Fasern verteilt sind und jeder Fasertyp die LD-Dynamik unterschiedlich reguliert, muss die Untersuchung von IMCLs fasertypspezifischsein 16,25,26,27. Daher verwendet dieses Protokoll Immunfluoreszenz auf seriellen Abschnitten, um Myosin Heavy Chain (MyHC) Isoform(en) zu identifizieren, die von jeder Faser exprimiert werden. Ein weiterer Vorteil dieses Protokolls ist die gleichzeitige Verarbeitung eines glykolytischen (Extensor digitorum longus, EDL) und eines oxidativen (Soleus) Muskels, die vor dem Einfrieren nebeneinander angeordnet sind (Abbildung 1). Diese gleichzeitige Verarbeitung spart nicht nur Zeit, sondern vermeidet auch Variabilität durch die getrennte Verarbeitung der Proben.
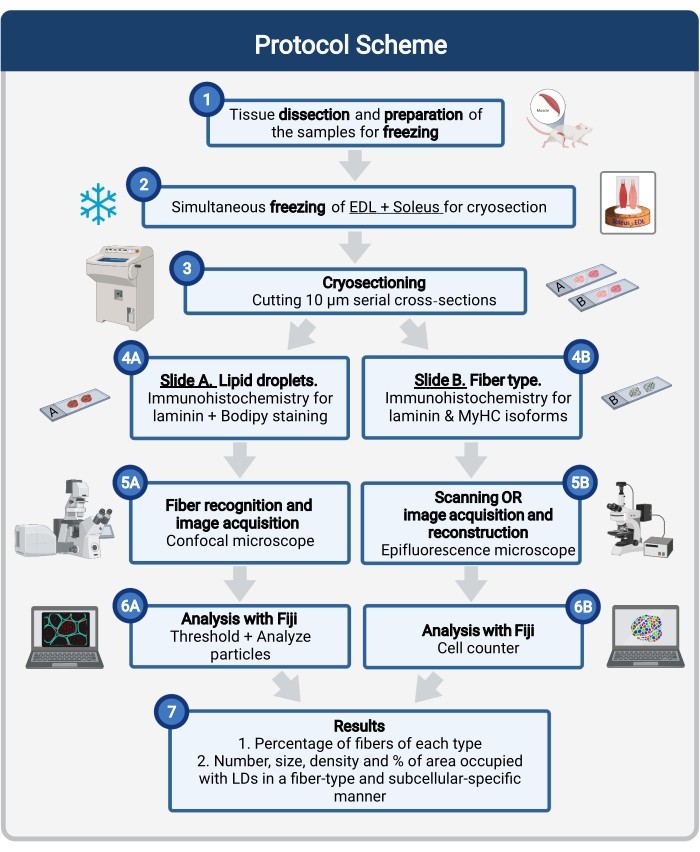
Abbildung 1: Schematische Übersicht des Vorgehens. Nach der Muskeldissektion (1) werden ausgewählte Muskeln ähnlicher Größe vorbereitet und zusammen eingefroren (2). Serielle Querschnitte von 10 μm werden mit einem Kryostaten erhalten und direkt auf Adhäsionsobjektträgern montiert (3). Aus zwei seriellen Objektträgern ist der erste (4A) für Laminin immunmarkiert und mit Bodipy gefärbt, um LDs zu erkennen, und der zweite (4B) ist mit Antikörpern gegen MyHCs zur Erkennung von Muskelfasertypen immunresistent. Die Bilder werden mit einem konfokalen Mikroskop für Bodipy (5A) und einem Epifluoreszenzmikroskop für Muskelfasertypen (5B) aufgenommen. Die Bilder werden in Fidschi analysiert, indem ein Schwellenwert angewendet und Partikel (6A) quantifiziert werden, um die Anzahl, die durchschnittliche Größe, die Dichte und den Prozentsatz der Gesamtfläche zu erhalten, die von LDs (7) oder Zählzellen (6B) eingenommen wird, um den Prozentsatz der Fasern jedes Typs im Abschnitt (7) zu erhalten. Abkürzungen: LDs = Lipidtröpfchen; EDL = Extensor digitorum longus; MyHCs = Myosin-Schwerkettenisoformen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
