חדירת שומנים בשריר השלד, המכונה מיואסטאטוזיס, עולה עם השמנת יתר והזדקנות. מיואסטאטוזיס מתואמת באופן שלילי עם מסת שריר וכוח ועם רגישות לאינסולין1. יתר על כן, מחקרים אחרונים מצביעים על כך שמידת המיוסטאטוזיס יכולה לשמש כגורם פרוגנוסטי למצבים אחרים כגון מחלות לב וכלי דם2, מחלת כבד שומני שאינה אלכוהולית3, או סרטן4. שומנים יכולים להצטבר בשרירי השלד בין סיבי השריר כשומנים חוץ-תאיים או בתוך הסיבים, כמו שומנים תוך-תאיים (IMCLs). IMCLs מאוחסנים בעיקר כטריגליצרידים בטיפות שומנים (LDs) המשמשים כדלק מטבולי במהלך פעילות גופנית 5,6. עם זאת, כאשר היצע השומנים עולה על הביקוש, או כאשר המיטוכונדריה הופכת לבלתי מתפקדת, IMCLs יהיו מעורבים בתנגודת לאינסולין בשרירים, כפי שניתן לראות אצל אנשים לא בריאים מבחינה מטבולית, שמנים ובחולי סוכרת מסוג 27. באופן מסקרן, לספורטאי סיבולת יש רמות דומות, אם לא גבוהות יותר, של IMCLs לאלה שנמצאו בחולים שמנים עם סוכרת מסוג 2, תוך שמירה על רגישות גבוהה לאינסולין. תופעה זו מתוארת כ”פרדוקס הספורטאי”8,9, והיא מוסברת על ידי הערכה ניואנסית יותר של LDs שרירים, הקשורים לגודלם, צפיפותם, לוקליזציה, דינמיקה והרכב מיני שומנים.
ראשית, גודל LD נמצא בקורלציה הפוכה לרגישות לאינסולין ולכושר גופני10,11. למעשה, LDs קטנים יותר מציגים שטח פנים גדול יחסית לפעולת ליפאז, ולכן, יש להם יכולת גדולה יותר לגייס שומנים12. שנית, צפיפות LD (מספר/משטח) ממלאת תפקיד שנוי במחלוקת בפעולתהאינסולין 8,10; עם זאת, נראה כי הוא גדל בספורטאים. שלישית, הלוקליזציה התת-תאית של LDs חשובה, שכן LDs הממוקמים ממש מתחת לממברנה של פני השטח (תת-סרקולמל או היקפי) מפעילים השפעה מזיקה יותר על הרגישות לאינסולין מאשר אלה המרכזיים 8,9,13. אלה האחרונים מספקים דלק למיטוכונדריה מרכזית, שיש לה פעילות נשימתית גדולה יותר ומתמחה יותר כדי לענות על הביקוש האנרגטי הגבוה הנדרש להתכווצות14. לעומת זאת, LDs היקפיים מספקים מיטוכונדריה תת-סרקולמלית, המעורבים בתהליכים הקשורים לממברנה8. לבסוף, מעבר לטריגליצרידים, שומנים מורכבים ספציפיים בתוך השריר עשויים להיות מזיקים יותר מאחרים. לדוגמה, דיאצילגליצרול, אציל-CoA ארוך שרשרת וסרמידים עשויים להצטבר בשריר כאשר שיעור תחלופת הטריגליצרידים נמוך, ובכך לפגוע באינסולין המאותתעל 9,15. אם נחזור ל”פרדוקס של הספורטאי”, לספורטאי סיבולת יש מספר גבוה של LDs מרכזיים קטנים יותר עם שיעורי תחלופה גבוהים בסיבים מסוג I (חמצוניים), בעוד שלחולי השמנת יתר וסוכרת יש LD היקפיים גדולים יותר עם שיעורי תחלופה נמוכים יותר בסיבים מסוג II (גליקוליטי) 8,15,16. בנוסף לתפקידם באגירת ושחרור אנרגיה, LDs באמצעות חומצות שומן נגזרות (FA) וחלבון פרווה (פריליפין 5) יכולים גם לתפקד כשחקנים קריטיים המעורבים בוויסות השעתוק של חמצון FA וביוגנזה מיטוכונדריאלית8. בגלל ההשלכות המכריעות שלהם בפיזיולוגיה ובפתופיזיולוגיה, יש הצדקה למחקרים מעמיקים על דינמיקה ותפקודים של LDs.
למרות שישנן מספר טכניקות לחקר IMCLs, לא כולן מתאימות לכימות מדויק של גודל LD, צפיפותו והתפלגותו באופן ספציפי לסיבים. לדוגמה, ההערכה של IMCLs על ידי ספקטרוסקופיית תהודה מגנטית, למרות היותה לא פולשנית, מציעה רמת רזולוציה שאינה מספיקה כדי לחקור את הגודל והמיקום המדויק של LDs בתוך הסיבים, והיא אינה ספציפית מסוג סיבים17,18. כמו כן, טכניקות ביוכימיות המבוצעות על הומוגנטים של שרירים שלמים19 אינן יכולות להעריך את מיקומם וגודלם של שומנים. כתוצאה מכך, השיטה המתאימה ביותר לניתוח המורפולוגיה והמיקום של LD היא מיקרוסקופיה אלקטרונית של שידור כמותי13, אך טכניקה זו יקרה וגוזלת זמן רב. לכן, הדמיה פלואורסצנטית קונפוקלית על תכשירים עם צבעים כגון שמן אדום O (ORO)20,21, monodansylpentane (MDH)22, או Bodipy 23,24,25, התגלתה ככלי הטוב ביותר למחקרים אלה.
כאן מתואר פרוטוקול שלם, הכולל דגימה ועיבוד של רקמות, צביעת Bodipy, ורכישה וניתוח של תמונה קונפוקלית כדי לכמת את הגודל, המספר והלוקליזציה של LD בקריואזות שרירי העכבר. מאחר ש-IMCLs אינם מחולקים באופן שווה בין סיבים חמצוניים וגליקוליטיים, וכל סוג סיבים מווסת את הדינמיקה של LD באופן שונה, המחקר של IMCLs חייב להיות ספציפי מסוג סיבים 16,25,26,27. לכן, פרוטוקול זה משתמש באימונופלואורסצנציה בחתכים סדרתיים כדי לזהות איזופורמים של שרשרת כבדת מיוזין (MyHC) המבוטאים על ידי כל סיב. יתרון נוסף של פרוטוקול זה הוא עיבוד סימולטני של שריר גליקוליטי (extensor digitorum longus, EDL) ושריר חמצוני (סולוס) המונחים זה לצד זה לפני הקפאה (איור 1). עיבוד סימולטני זה לא רק חוסך זמן אלא גם מונע השתנות עקב עיבוד נפרד של הדגימות.
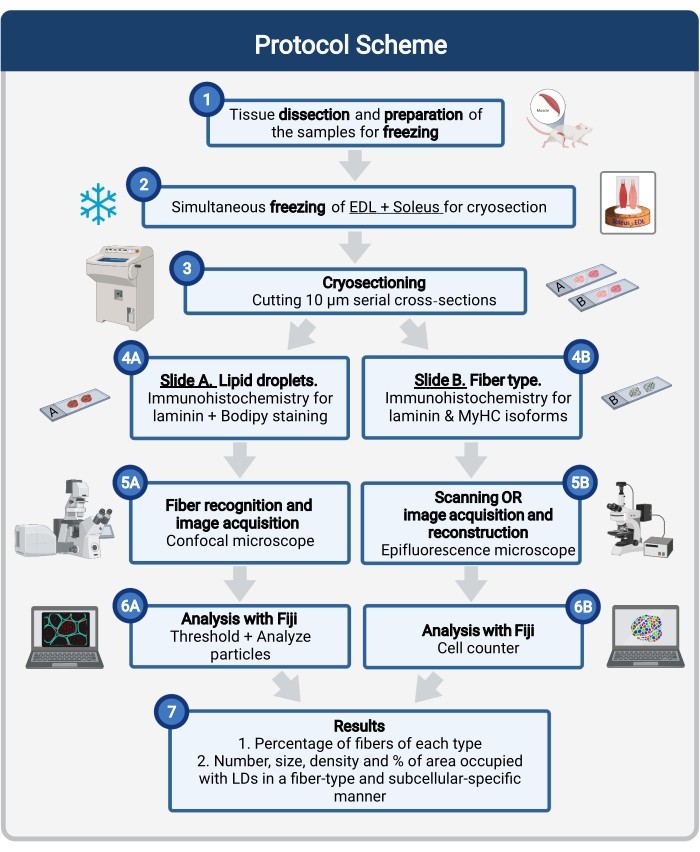
איור 1: סקירה סכמטית של ההליך. לאחר כריתת שרירים (1), השרירים שנבחרו בגודל דומה מוכנים ומקפואים יחד (2). מקטעים רוחביים טוריים של 10 מיקרומטר מתקבלים באמצעות קריוסטאט ומורכבים ישירות על שקופיות הידבקות (3). משתי שקופיות סדרתיות, הראשונה (4A) מסומנת באופן חיסוני ללאמינין ומוכתמת ב-Bodipy כדי לזהות LDs והשנייה (4B) מוכתמת בנוגדנים נגד MyHCs לזיהוי סוגי סיבי שריר. התמונות נרכשות באמצעות מיקרוסקופ קונפוקלי עבור Bodipy (5A) ומיקרוסקופ אפיפלואורסצנטי לסוגי סיבי שריר (5B). התמונות מנותחות בפיג’י על ידי החלת סף וכימות חלקיקים (6A) כדי לקבל את המספר, הגודל הממוצע, הצפיפות והאחוז של השטח הכולל שנכבש על ידי LDs (7) או ספירת תאים (6B) כדי לקבל את אחוז הסיבים מכל סוג בסעיף (7). קיצורים: LDs = טיפות שומנים; EDL = extensor digitorum longus; MyHCs = איזופורמים של שרשרת כבדה של מיוזין. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
